

Endosymbiont or host: who drove mitochondrial and plastid evolution?
- PMID: 21333023
- PMCID: PMC3050876
- DOI: 10.1186/1745-6150-6-12
Endosymbiont or host: who drove mitochondrial and plastid evolution?
Abstract
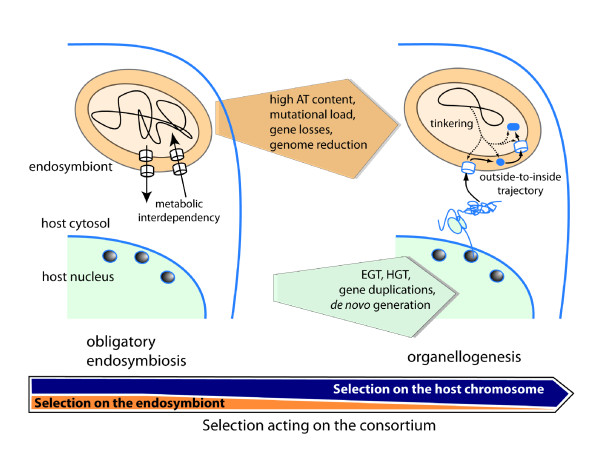
The recognition that mitochondria and plastids are derived from alphaproteobacterial and cyanobacterial endosymbionts, respectively, was one of the greatest advances in modern evolutionary biology. Researchers have yet however to provide detailed cell biological descriptions of how these once free-living prokaryotes were transformed into intracellular organelles. A key area of study in this realm is elucidating the evolution of the molecular machines that control organelle protein topogenesis. Alcock et al. (Science 2010, 327 [5966]:649-650) suggest that evolutionary innovations that established the mitochondrial protein sorting system were driven by the alphaproteobacterial endosymbiont (an "insiders' perspective"). In contrast, here we argue that evolution of mitochondrial and plastid topogenesis may better be understood as an outcome of selective pressures acting on host cell chromosomes (the "outsiders' view").
Figures
Similar articles
-
How do endosymbionts become organelles? Understanding early events in plastid evolution.Bioessays. 2007 Dec;29(12):1239-46. doi: 10.1002/bies.20671. Bioessays. 2007. PMID: 18027391
-
Origin and evolution of organelle genomes.Curr Opin Genet Dev. 1993 Dec;3(6):884-90. doi: 10.1016/0959-437x(93)90009-e. Curr Opin Genet Dev. 1993. PMID: 8118213 Review.
-
Mitochondrial and plastid evolution in eukaryotes: an outsiders' perspective.Nat Rev Genet. 2009 Jul;10(7):495-505. doi: 10.1038/nrg2610. Nat Rev Genet. 2009. PMID: 19506574 Review.
-
Dinoflagellates with relic endosymbiont nuclei as models for elucidating organellogenesis.Proc Natl Acad Sci U S A. 2020 Mar 10;117(10):5364-5375. doi: 10.1073/pnas.1911884117. Epub 2020 Feb 24. Proc Natl Acad Sci U S A. 2020. PMID: 32094181 Free PMC article.
-
Retrotransposition facilitated the establishment of a primary plastid in the thecate amoeba Paulinella.Proc Natl Acad Sci U S A. 2022 Jun 7;119(23):e2121241119. doi: 10.1073/pnas.2121241119. Epub 2022 May 31. Proc Natl Acad Sci U S A. 2022. PMID: 35639693 Free PMC article.
Cited by
-
Protein import into the photosynthetic organelles of Paulinella chromatophora and its implications for primary plastid endosymbiosis.Symbiosis. 2012 Dec;58(1-3):99-107. doi: 10.1007/s13199-012-0202-2. Epub 2012 Dec 13. Symbiosis. 2012. PMID: 23482692 Free PMC article.
-
Protein import into bacterial endosymbionts and evolving organelles.FEBS J. 2025 Jun;292(12):2992-3013. doi: 10.1111/febs.17356. Epub 2024 Dec 10. FEBS J. 2025. PMID: 39658314 Free PMC article. Review.
-
Cytochrome c oxidase: evolution of control via nuclear subunit addition.Biochim Biophys Acta. 2012 Apr;1817(4):590-7. doi: 10.1016/j.bbabio.2011.07.007. Epub 2011 Jul 23. Biochim Biophys Acta. 2012. PMID: 21802404 Free PMC article. Review.
-
Improved plant biomass production under low nitrogen conditions through conditional accumulation of the second messenger, guanosine tetraphosphate, in chloroplasts and mitochondria.Front Plant Sci. 2025 Jan 13;15:1524665. doi: 10.3389/fpls.2024.1524665. eCollection 2024. Front Plant Sci. 2025. PMID: 39872205 Free PMC article.
-
Cross-Species Functional Conservation and Possible Origin of the N-Terminal Specificity Domain of Mitochondrial Presequences.Front Plant Sci. 2020 Feb 13;11:64. doi: 10.3389/fpls.2020.00064. eCollection 2020. Front Plant Sci. 2020. PMID: 32117399 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
