

Color in the cortex: single- and double-opponent cells
- PMID: 21333672
- PMCID: PMC3121536
- DOI: 10.1016/j.visres.2011.02.012
Color in the cortex: single- and double-opponent cells
Abstract
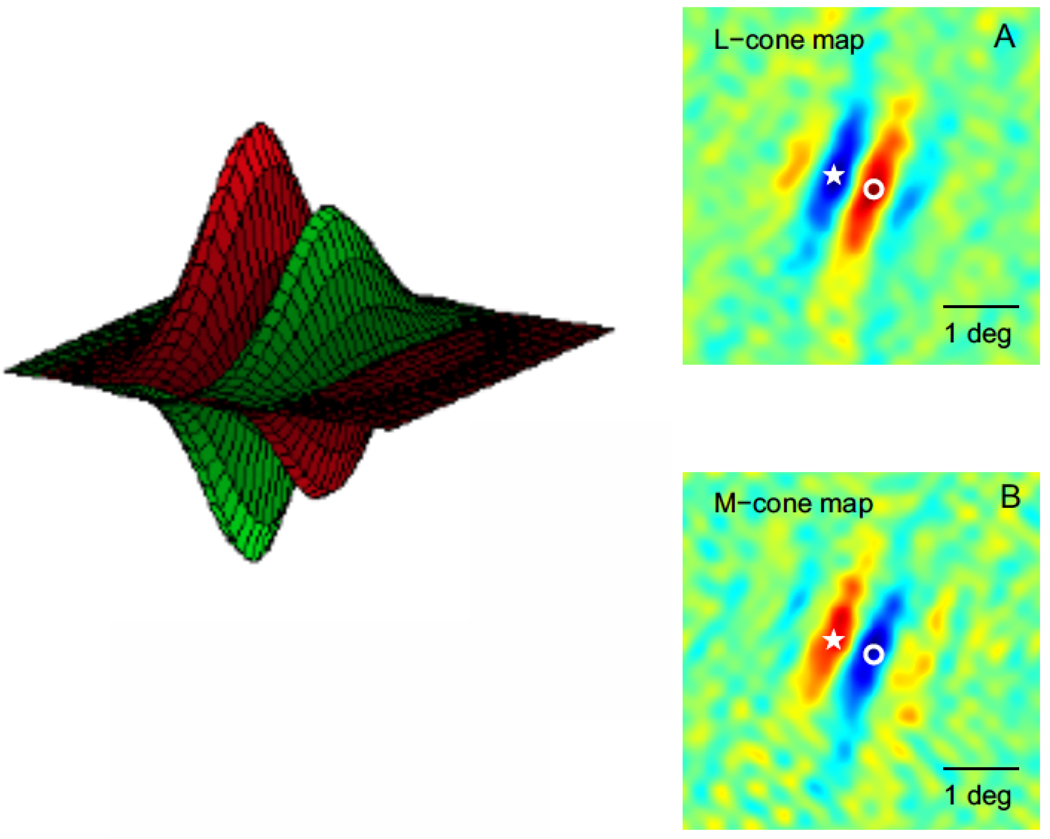
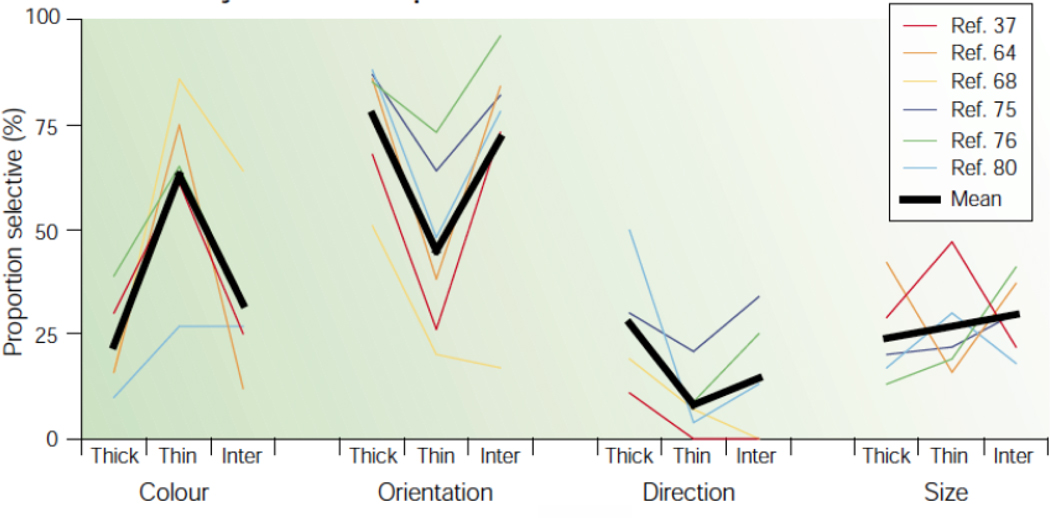
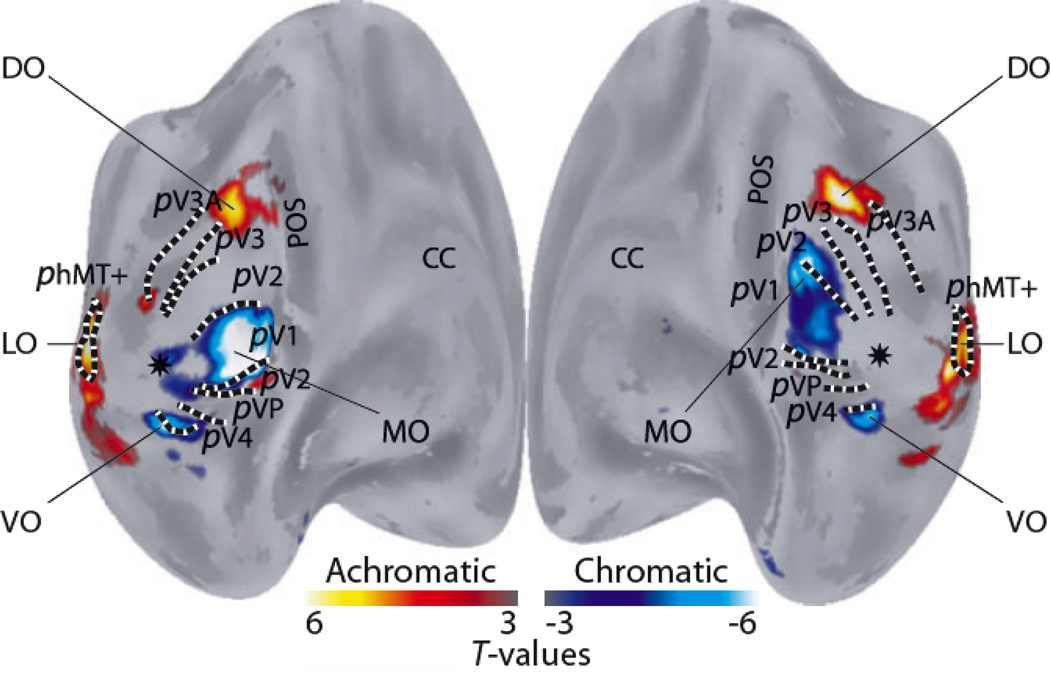
This is a review of the research during the past 25years on cortical processing of color signals. At the beginning of the period the modular view of cortical processing predominated. However, at present an alternative view, that color and form are linked inextricably in visual cortical processing, is more persuasive than it seemed in 1985. Also, the role of the primary visual cortex, V1, in color processing now seems much larger than it did in 1985. The re-evaluation of the important role of V1 in color vision was caused in part by investigations of human V1 responses to color, measured with functional magnetic resonance imaging, fMRI, and in part by the results of numerous studies of single-unit neurophysiology in non-human primates. The neurophysiological results have highlighted the importance of double-opponent cells in V1. Another new concept is population coding of hue, saturation, and brightness in cortical neuronal population activity.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures

References
-
- Bartels A, Zeki S. The architecture of the colour centre in the human visual brain: new results and a review. Eur. J. Neurosci. 2000;12:172–193. - PubMed
-
- Beauchamp MS, Haxby JV, Jennings JE, DeYoe EA. An fMRI version of the Farnsworth-Munsell 100-Hue test reveals multi- ple color-selective areas in human ventral occipitotemporal cortex. Cereb. Cortex. 1999;9:257–263. - PubMed
-
- Beaudot WH, Mullen KT. Orientation selectivity in luminance and color vision assessed using 2-d band-pass filtered spatial noise. Vision Res. 2005;45:687–696. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
