

The complex logic of stringent response regulation in Caulobacter crescentus: starvation signalling in an oligotrophic environment
- PMID: 21338423
- PMCID: PMC3093662
- DOI: 10.1111/j.1365-2958.2011.07602.x
The complex logic of stringent response regulation in Caulobacter crescentus: starvation signalling in an oligotrophic environment
Abstract
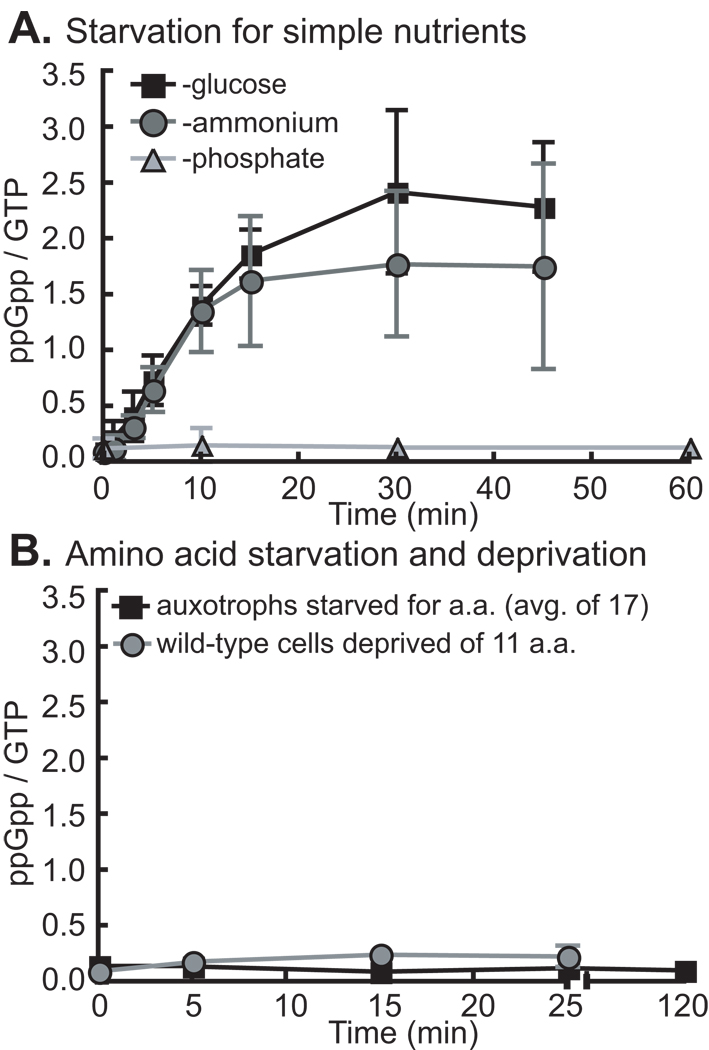
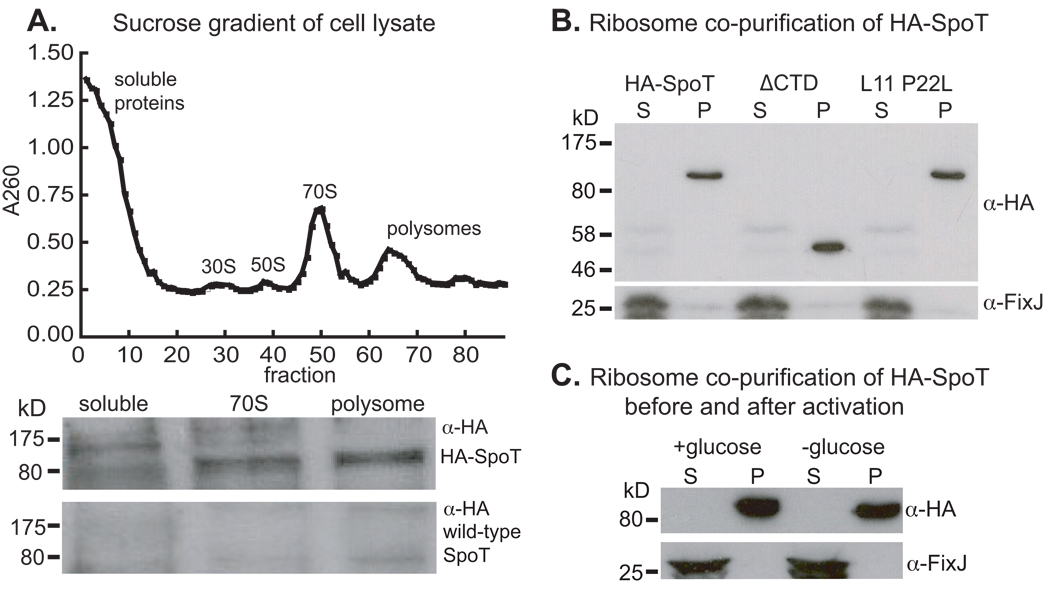
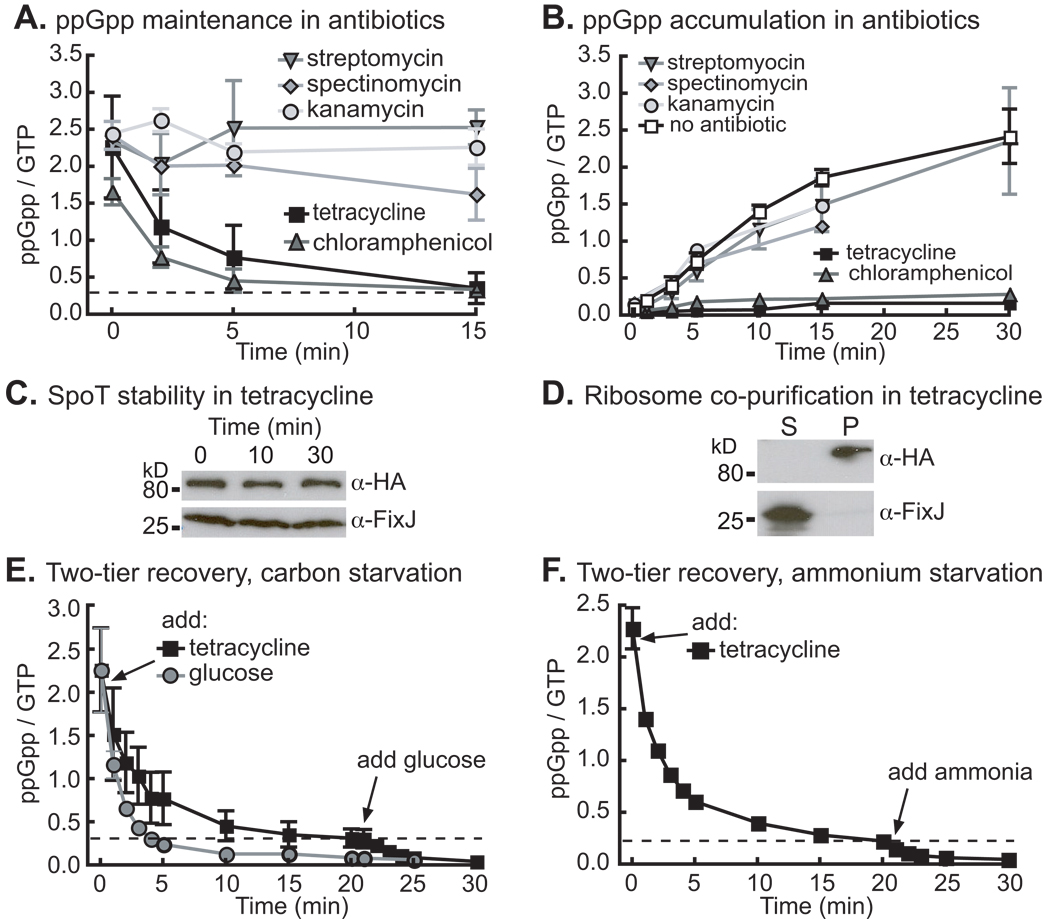
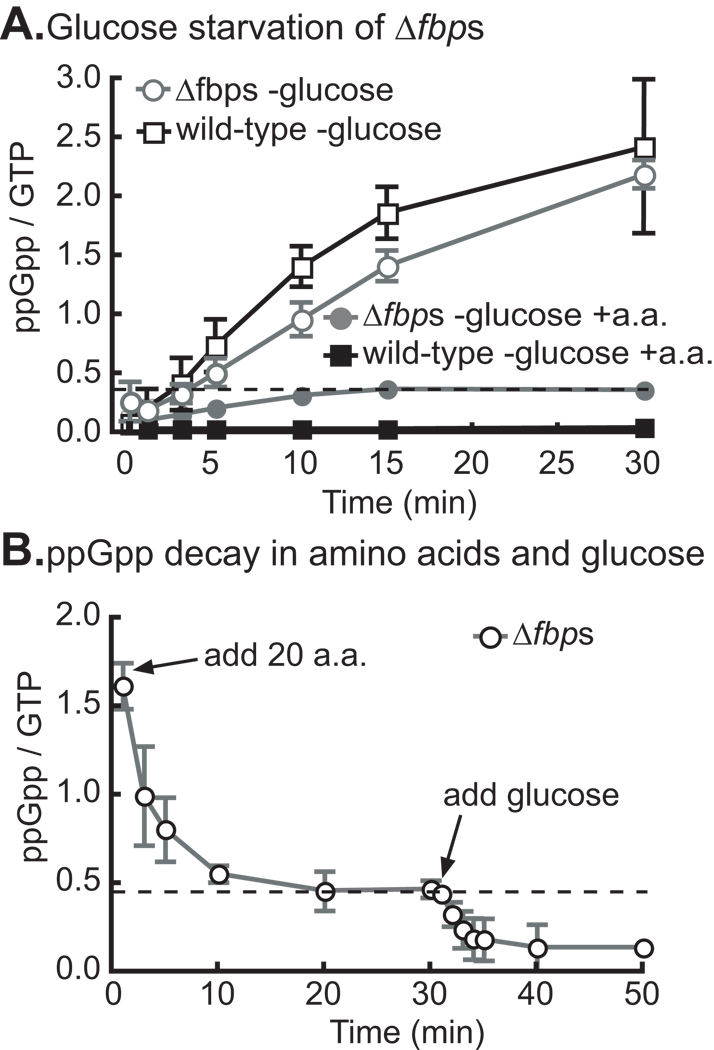
Bacteria rapidly adapt to nutritional changes via the stringent response, which entails starvation-induced synthesis of the small molecule, ppGpp, by RelA/SpoT homologue (Rsh) enzymes. Binding of ppGpp to RNA polymerase modulates the transcription of hundreds of genes and remodels the physiology of the cell. Studies of the stringent response have primarily focused on copiotrophic bacteria such as Escherichia coli; little is known about how stringent signalling is regulated in species that live in consistently nutrient-limited (i.e. oligotrophic) environments. Here we define the input logic and transcriptional output of the stringent response in the oligotroph, Caulobacter crescentus. The sole Rsh protein, SpoT(CC), binds to and is regulated by the ribosome, and exhibits AND-type control logic in which amino acid starvation is a necessary but insufficient signal for activation of ppGpp synthesis. While both glucose and ammonium starvation upregulate the synthesis of ppGpp, SpoT(CC) detects these starvation signals by two independent mechanisms. Although the logic of stringent response control in C. crescentus differs from E. coli, the global transcriptional effects of elevated ppGpp are similar, with the exception of 16S rRNA transcription, which is controlled independently of spoT(CC). This study highlights how the regulatory logic controlling the stringent response may be adapted to the nutritional niche of a bacterial species.
© 2011 Blackwell Publishing Ltd.
Figures
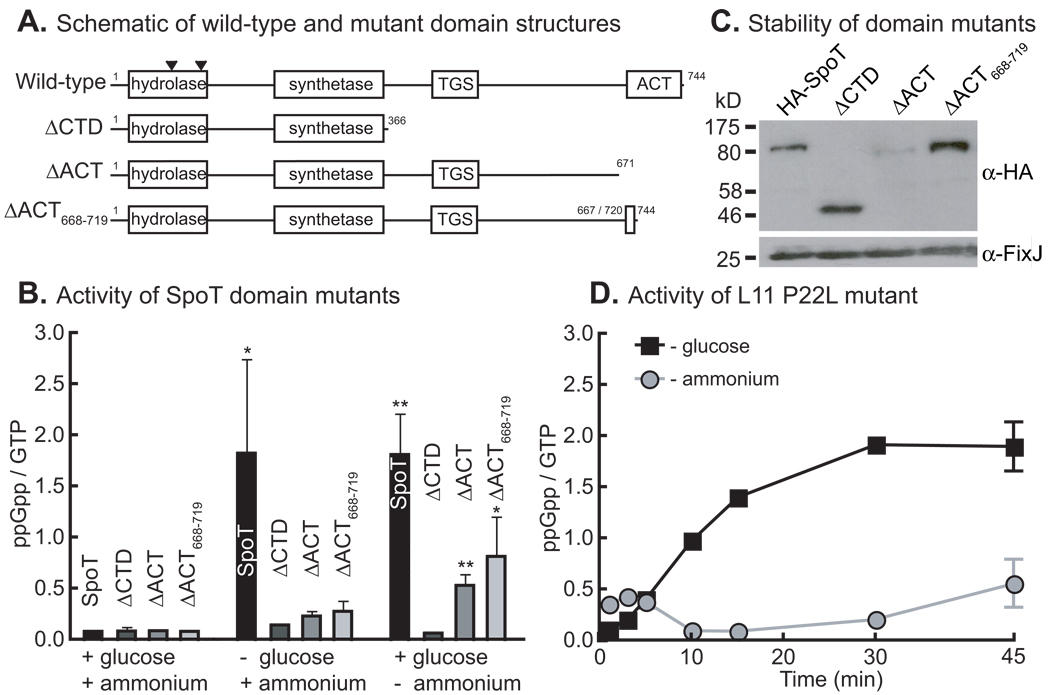
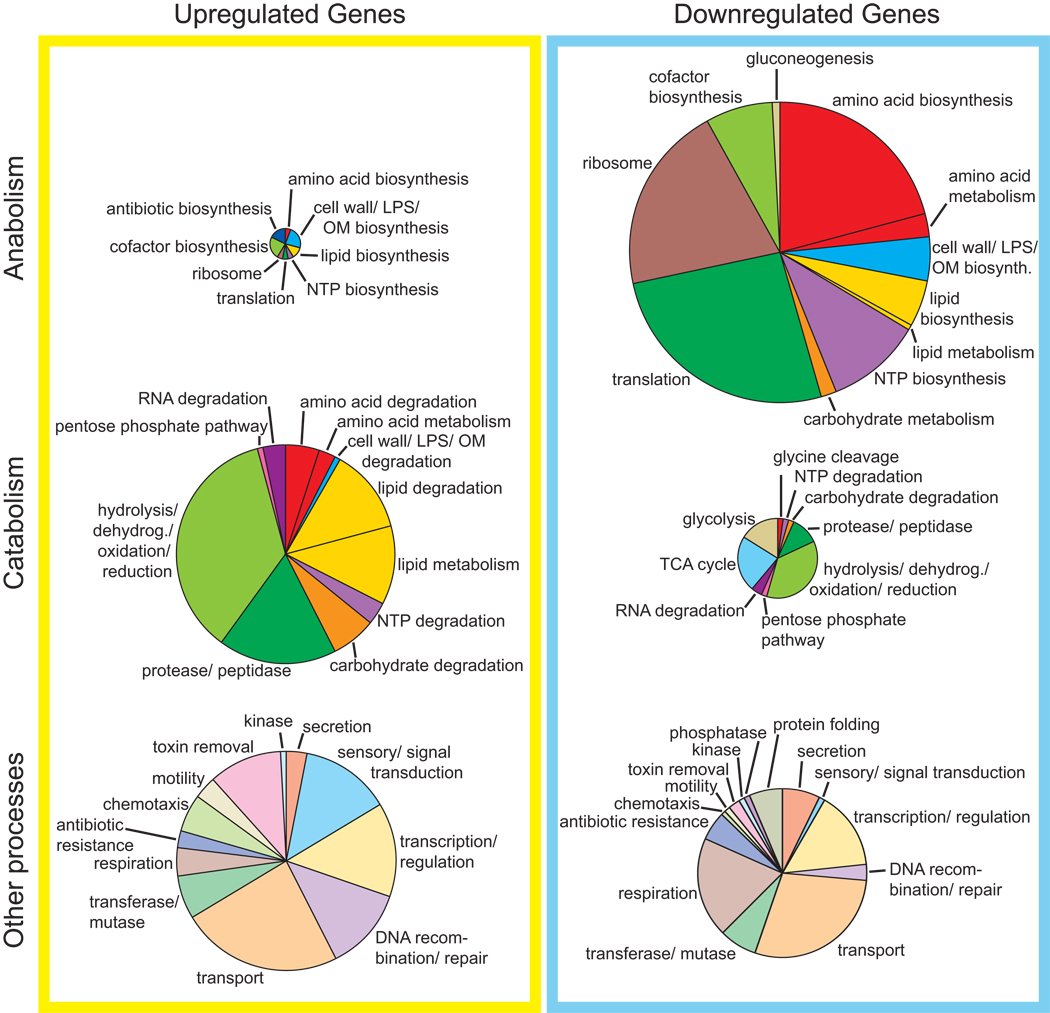
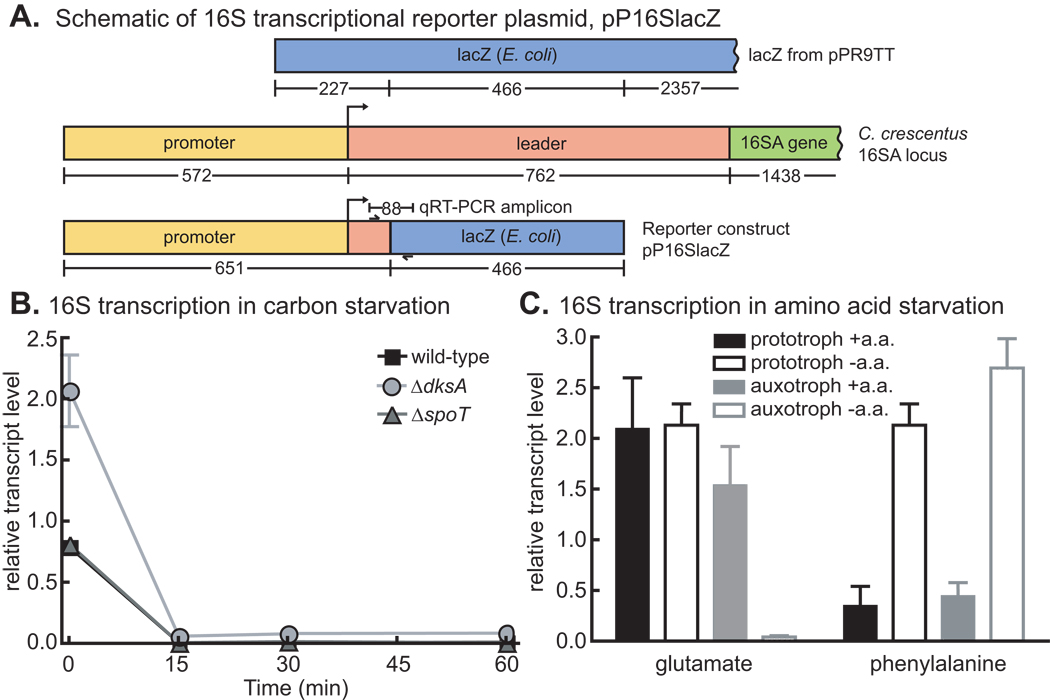
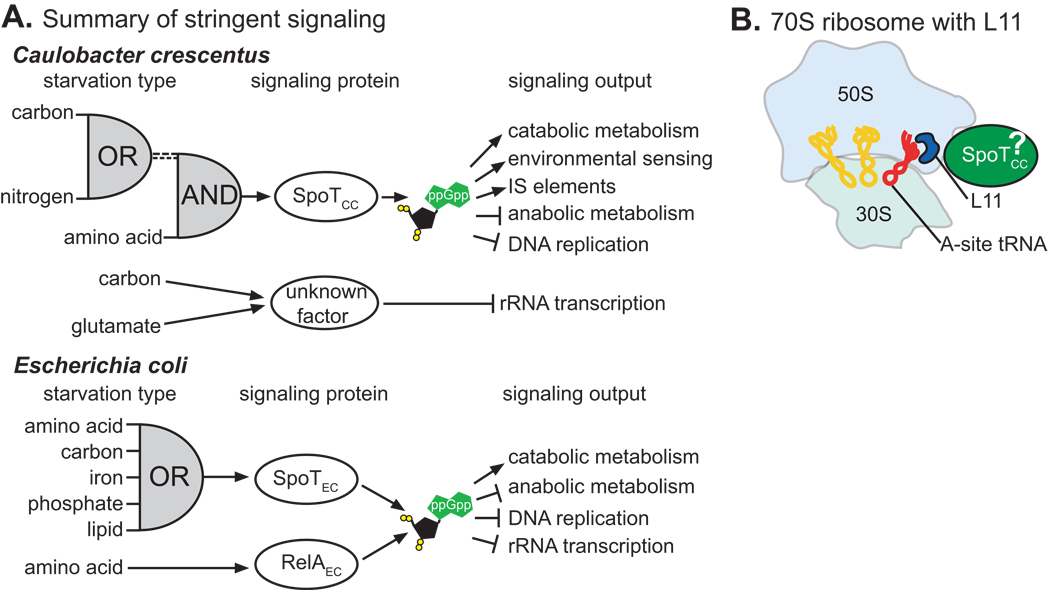
References
-
- Amemiya K. Conserved Sequence Elements Upstream and Downstream from the Transcription Initiation Site of the Caulobacter crescentus rrnA Gene Cluster. J Mol Biol. 1989;210:245–254. - PubMed
-
- Aravind L, Koonin EV. The HD domain defins a new superfamily of metal-depedent phosphohydrolases. Trends Biochem Sci. 1998;23:469–472. - PubMed
-
- Avarbock D, Avarbock A, Rubin H. Differential Regulation of Opposing RelMtb Activity by the Aminoacylation State of a tRNA.Ribosome.mRNA.RelMtb Complex. Biochemistry. 2000;39:11640–11648. - PubMed
-
- Avarbock D, Salem J, Li L, Wang Z, Rubin H. Cloning and characterization of a bifunctional RelA/SpoT homologue from Mycobacterium tuberculosis. Gene. 1999;233:261–269. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
