

Expression of the Distalless-B gene in Ciona is regulated by a pan-ectodermal enhancer module
- PMID: 21338600
- PMCID: PMC3081900
- DOI: 10.1016/j.ydbio.2011.02.009
Expression of the Distalless-B gene in Ciona is regulated by a pan-ectodermal enhancer module
Abstract
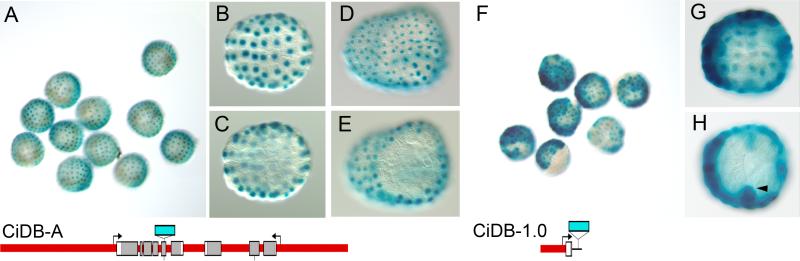
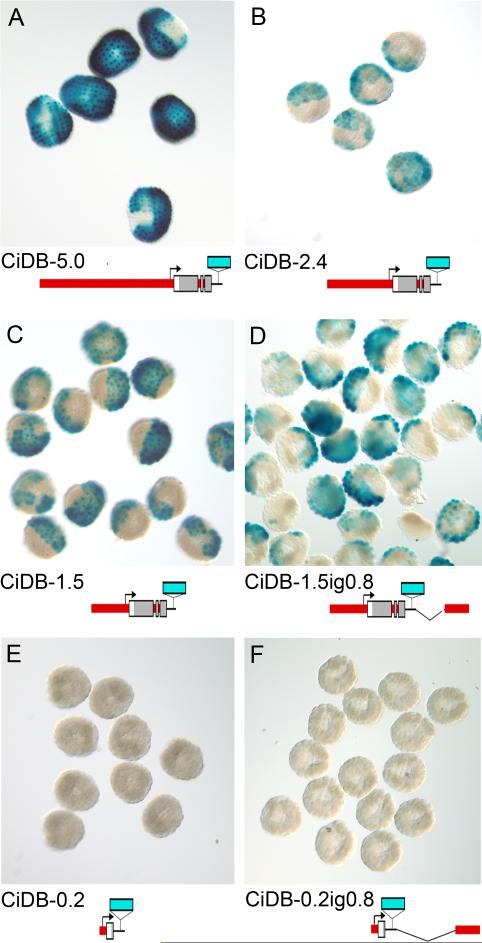
The Ci-Dll-B gene is an early regulator of ectodermal development in the ascidian Ciona intestinalis (Imai et al., 2006). Ci-Dll-B is located in a convergently transcribed bigene cluster with a tandem duplicate, Ci-Dll-A. This clustered genomic arrangement is the same as those of the homologous vertebrate Dlx genes, which are also arranged in convergently transcribed bigene clusters. Sequence analysis of the C. intestinalis Dll-A-B cluster reveals a 378bp region upstream of Ci-Dll-B, termed B1, which is highly conserved with the corresponding region from the congener Ciona savignyi. The B1 element is necessary and sufficient to drive expression of a lacZ reporter gene in a pattern mimicking the endogenous expression of Ci-Dll-B at gastrula stages. This expression pattern which is specific to the entire animal hemisphere is activated preferentially in posterior, or b-lineage, cells by a central portion of B1. Expression in anterior, or a-lineage cells, can be activated by this central portion in combination with the distal part of B1. Anterior expression can also be activated by the central part of B1 plus both the proximal part of B1 and non-conserved sequence upstream of B1. Thus, cis-regulation of early Ci-Dll-B expression is activated by a required submodule in the center of B1, driving posterior expression, which works in combination with redundant submodules that respond to differentially localized anterior factors to produce the total animal hemisphere expression pattern. Interestingly, the intergenic region of the cluster, which is important for expression of the Dlx genes in vertebrates, does not have a specific activating function in the reporter genes tested, but acts as an attenuator in combination with upstream sequences.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures




References
-
- Caracciolo A, Gregorio AD, Aniello F, Lauro RD, Branno M. Identification and developmental expression of three Distal-less homeobox containing genes in the ascidian Ciona intestinalis. Mech. Dev. 2000;99:173–176. - PubMed
-
- Di Gregorio A, Corbo J, Levine M. The regulation of forkhead/HNf-3 beta expression in the Ciona embryo. Dev. Biol. 2001;229:31–43. - PubMed
-
- Ellies DL, Stock DW, Hatch G, Giroux G, Weiss KM, Ekker M. Relationship between the genomic organization and the overlapping embryonic expression patterns of the zebrafish dlx genes. Genomics. 1997;45:580–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
