

The frontotemporal dementia mutation R406W blocks tau's interaction with the membrane in an annexin A2-dependent manner
- PMID: 21339331
- PMCID: PMC3044115
- DOI: 10.1083/jcb.201007161
The frontotemporal dementia mutation R406W blocks tau's interaction with the membrane in an annexin A2-dependent manner
Abstract
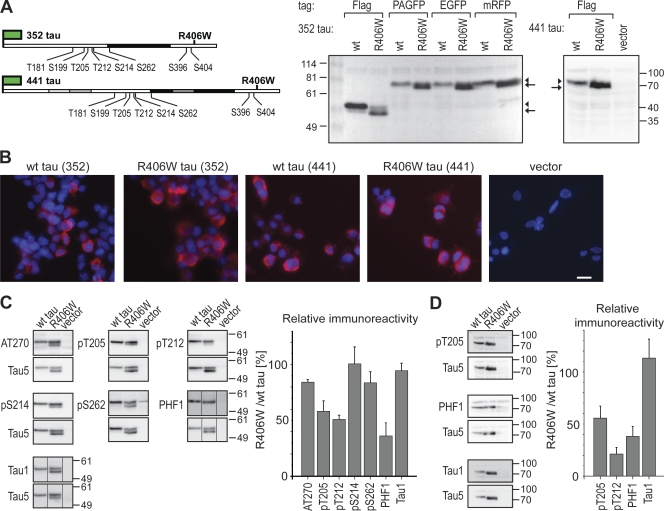
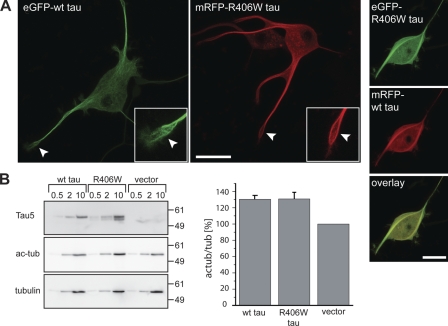
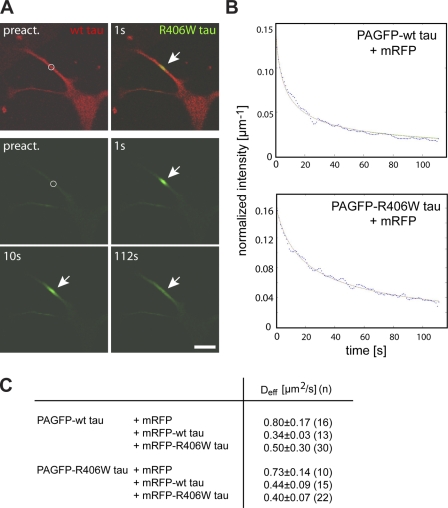
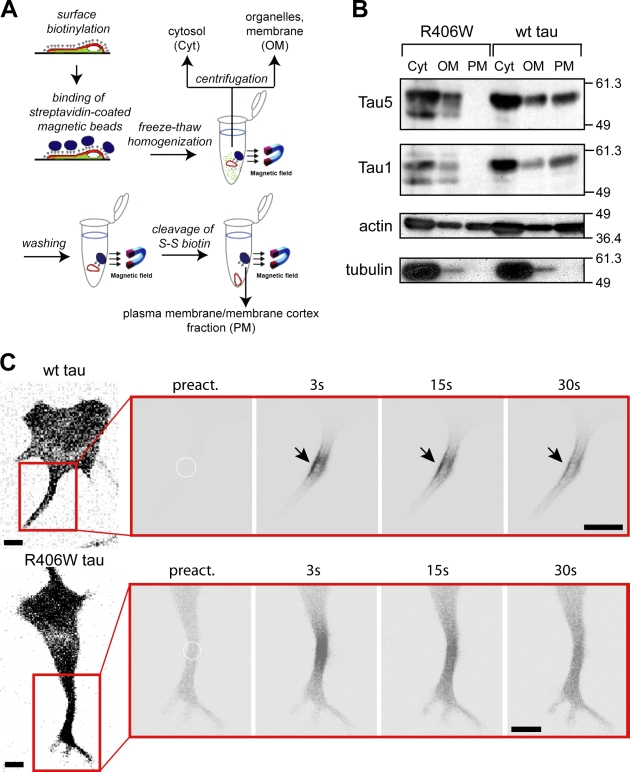
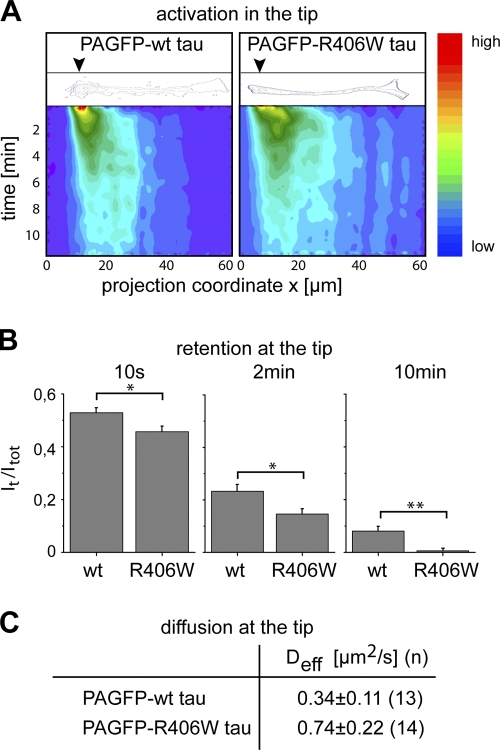
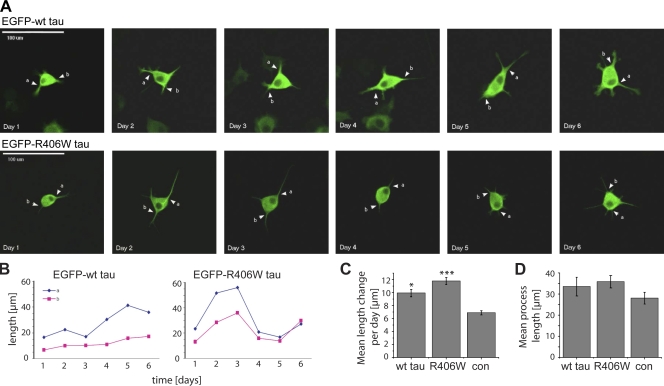
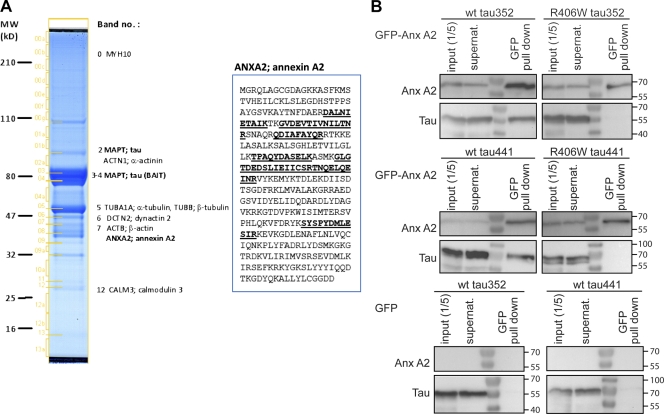
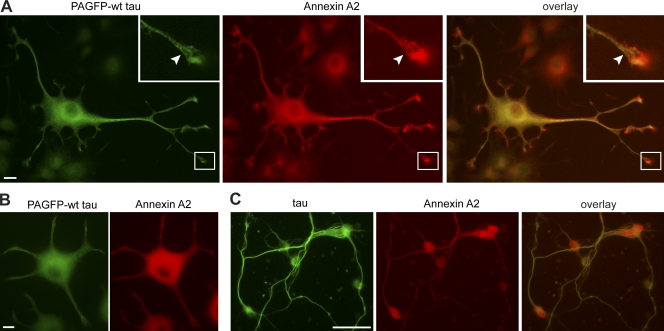
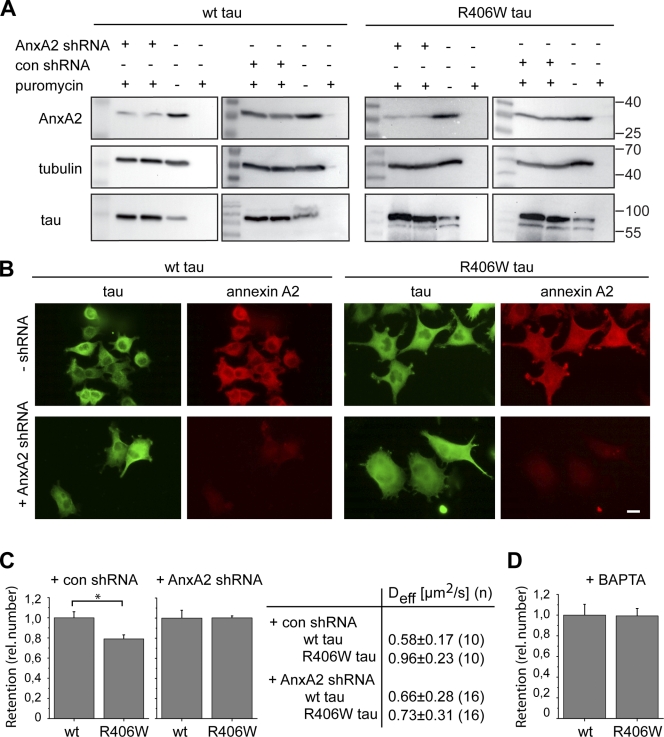
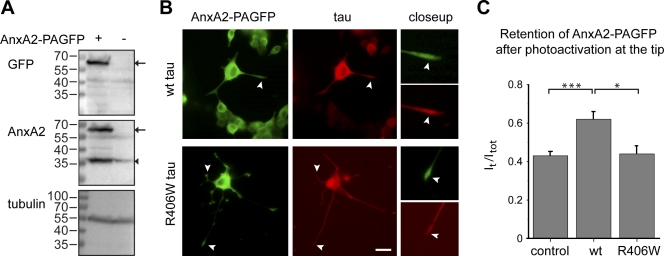
Changes of the microtubule-associated protein tau are central in Alzheimer's disease (AD) and frontotemporal dementia with Parkinsonism linked to chromosome 17 (FTDP-17). However, the functional consequence of the FTDP-17 tau mutation R406W, which causes a tauopathy clinically resembling AD, is not well understood. We find that the R406W mutation does not affect microtubule interaction but abolishes tau's membrane binding. Loss of binding is associated with decreased trapping at the tip of neurites and increased length fluctuations during process growth. Tandem affinity purification tag purification and mass spectrometry identify the calcium-regulated plasma membrane-binding protein annexin A2 (AnxA2) as a potential interaction partner of tau. Consistently, wild-type tau but not R406W tau interacts with AnxA2 in a heterologous yeast expression system. Sequestration of Ca(2+) or knockdown of AnxA2 abolishes the differential trapping of wild-type and R406W tau. We suggest that the pathological effect of the R406W mutation is caused by impaired membrane binding, which involves a functional interaction with AnxA2 as a membrane-cytoskeleton linker.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
