

Sustained and rapid chromosome movements are critical for chromosome pairing and meiotic progression in budding yeast
- PMID: 21339478
- PMCID: PMC3120158
- DOI: 10.1534/genetics.110.125575
Sustained and rapid chromosome movements are critical for chromosome pairing and meiotic progression in budding yeast
Abstract
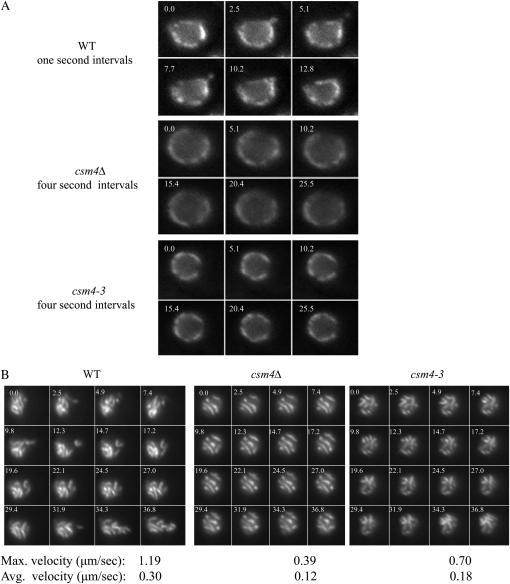
Telomere-led chromosome movements are a conserved feature of meiosis I (MI) prophase. Several roles have been proposed for such chromosome motion, including promoting homolog pairing and removing inappropriate chromosomal interactions. Here, we provide evidence in budding yeast that rapid chromosome movements affect homolog pairing and recombination. We found that csm4Δ strains, which are defective for telomere-led chromosome movements, show defects in homolog pairing as measured in a "one-dot/two-dot tetR-GFP" assay; however, pairing in csm4Δ eventually reaches near wild-type (WT) levels. Charged-to-alanine scanning mutagenesis of CSM4 yielded one allele, csm4-3, that confers a csm4Δ-like delay in meiotic prophase but promotes high spore viability. The meiotic delay in csm4-3 strains is essential for spore viability because a null mutation (rad17Δ) in the Rad17 checkpoint protein suppresses the delay but confers a severe spore viability defect. csm4-3 mutants show a general defect in chromosome motion but an intermediate defect in chromosome pairing. Chromosome velocity analysis in live cells showed that while average chromosome velocity was strongly reduced in csm4-3, chromosomes in this mutant displayed occasional rapid movements. Lastly, we observed that spo11 mutants displaying lower levels of meiosis-induced double-strand breaks showed higher spore viability in the presence of the csm4-3 mutation compared to csm4Δ. On the basis of these observations, we propose that during meiotic prophase the presence of occasional fast moving chromosomes over an extended period of time is sufficient to promote WT levels of recombination and high spore viability; however, sustained and rapid chromosome movements are required to prevent a checkpoint response and promote efficient meiotic progression.
Figures






Similar articles
-
Meiotic chromosome pairing is promoted by telomere-led chromosome movements independent of bouquet formation.PLoS Genet. 2012;8(5):e1002730. doi: 10.1371/journal.pgen.1002730. Epub 2012 May 24. PLoS Genet. 2012. PMID: 22654677 Free PMC article.
-
Csm4-dependent telomere movement on nuclear envelope promotes meiotic recombination.PLoS Genet. 2008 Sep 26;4(9):e1000196. doi: 10.1371/journal.pgen.1000196. PLoS Genet. 2008. PMID: 18818742 Free PMC article.
-
Rapid telomere movement in meiotic prophase is promoted by NDJ1, MPS3, and CSM4 and is modulated by recombination.Cell. 2008 Jun 27;133(7):1175-87. doi: 10.1016/j.cell.2008.04.047. Cell. 2008. PMID: 18585352
-
The nature of meiotic chromosome dynamics and recombination in budding yeast.J Microbiol. 2019 Apr;57(4):221-231. doi: 10.1007/s12275-019-8541-9. Epub 2019 Jan 22. J Microbiol. 2019. PMID: 30671743 Review.
-
Telomere-led meiotic chromosome movements: recent update in structure and function.Nucleus. 2020 Jan 1;11(9):111-116. doi: 10.1080/19491034.2020.1769456. Nucleus. 2020. PMID: 32412326 Free PMC article. Review.
Cited by
-
Meiotic chromosome pairing is promoted by telomere-led chromosome movements independent of bouquet formation.PLoS Genet. 2012;8(5):e1002730. doi: 10.1371/journal.pgen.1002730. Epub 2012 May 24. PLoS Genet. 2012. PMID: 22654677 Free PMC article.
-
Agent-based modeling of nuclear chromosome ensemble identifies determinants of homolog pairing during meiosis.bioRxiv [Preprint]. 2024 Jan 13:2023.08.09.552574. doi: 10.1101/2023.08.09.552574. bioRxiv. 2024. Update in: PLoS Comput Biol. 2024 May 13;20(5):e1011416. doi: 10.1371/journal.pcbi.1011416. PMID: 38260664 Free PMC article. Updated. Preprint.
-
Diffusion and distal linkages govern interchromosomal dynamics during meiotic prophase.Proc Natl Acad Sci U S A. 2022 Mar 22;119(12):e2115883119. doi: 10.1073/pnas.2115883119. Epub 2022 Mar 18. Proc Natl Acad Sci U S A. 2022. PMID: 35302885 Free PMC article.
-
Mixing and Matching Chromosomes during Female Meiosis.Cells. 2020 Mar 12;9(3):696. doi: 10.3390/cells9030696. Cells. 2020. PMID: 32178277 Free PMC article. Review.
-
Interorganelle interactions and inheritance patterns of nuclei and vacuoles in budding yeast meiosis.Autophagy. 2014 Feb;10(2):285-95. doi: 10.4161/auto.27192. Epub 2013 Dec 4. Autophagy. 2014. PMID: 24345927 Free PMC article.
References
-
- Alexandru G., Uhlmann F., Mechtler K., Poupart M. A., Nasmyth K., 2001. Phosphorylation of the cohesin subunit Scc1 by Polo/Cdc5 kinase regulates sister chromatid separation in yeast. Cell 105: 459–472 - PubMed
-
- Allers T., Lichten M., 2001. Intermediates of yeast meiotic recombination contain heteroduplex DNA. Mol. Cell 8: 225–231 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
