

In glycine and GABA(A) channels, different subunits contribute asymmetrically to channel conductance via residues in the extracellular domain
- PMID: 21343294
- PMCID: PMC3075687
- DOI: 10.1074/jbc.M110.204610
In glycine and GABA(A) channels, different subunits contribute asymmetrically to channel conductance via residues in the extracellular domain
Abstract
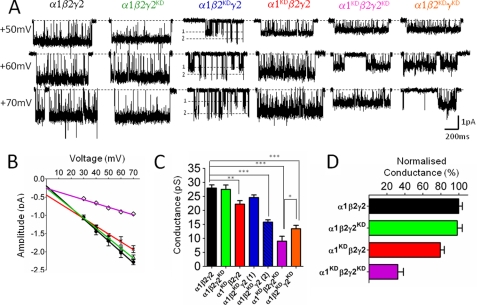
Single-channel conductance in Cys-loop channels is controlled by the nature of the amino acids in the narrowest parts of the ion conduction pathway, namely the second transmembrane domain (M2) and the intracellular helix. In cationic channels, such as Torpedo ACh nicotinic receptors, conductance is increased by negatively charged residues exposed to the extracellular vestibule. We now show that positively charged residues at the same loop 5 position boost also the conductance of anionic Cys-loop channels, such as glycine (α1 and α1β) and GABA(A) (α1β2γ2) receptors. Charge reversal mutations here produce a greater decrease on outward conductance, but their effect strongly depends on which subunit carries the mutation. In the glycine α1β receptor, replacing Lys with Glu in α1 reduces single-channel conductance by 41%, but has no effect in the β subunit. By expressing concatameric receptors with constrained stoichiometry, we show that this asymmetry is not explained by the subunit copy number. A similar pattern is observed in the α1β2γ2 GABA(A) receptor, where only mutations in α1 or β2 decreased conductance (to different extents). In both glycine and GABA receptors, the effect of mutations in different subunits does not sum linearly: mutations that had no detectable effect in isolation did enhance the effect of mutations carried by other subunits. As in the nicotinic receptor, charged residues in the extracellular vestibule of anionic Cys-loop channels influence elementary conductance. The size of this effect strongly depends on the direction of the ion flow and, unexpectedly, on the nature of the subunit that carries the residue.
Figures
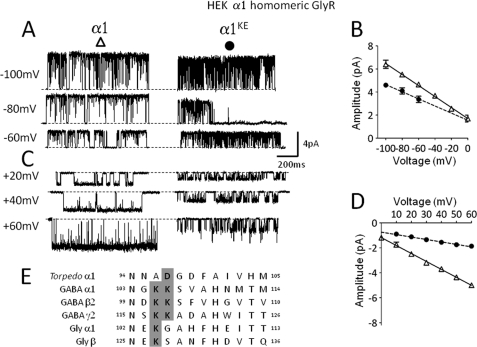
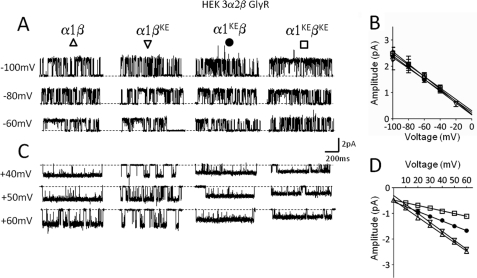
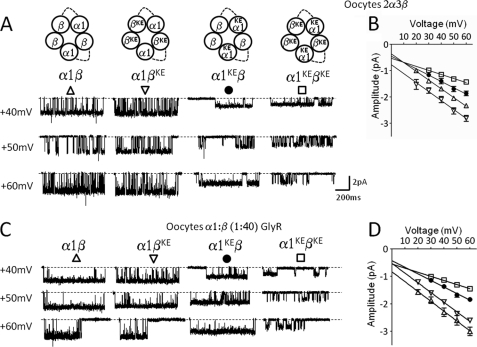
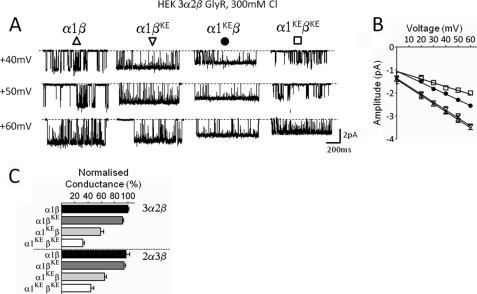
References
-
- Unwin N. (2005) J. Mol. Biol. 346, 967–989 - PubMed
-
- Hilf R. J., Dutzler R. (2009) Nature 457, 115–118 - PubMed
-
- Bocquet N., Nury H., Baaden M., Le, Poupon C., Changeux J. P., Delarue M., Corringer P. J. (2009) Nature 457, 111–114 - PubMed
-
- Imoto K., Busch C., Sakmann B., Mishina M., Konno T., Nakai J., Bujo H., Mori Y., Fukuda K., Numa S. (1988) Nature 335, 645–648 - PubMed
-
- Kelley S. P., Dunlop J. I., Kirkness E. F., Lambert J. J., Peters J. A. (2003) Nature 424, 321–324 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
