

Pathways for Holliday junction processing during homologous recombination in Saccharomyces cerevisiae
- PMID: 21343337
- PMCID: PMC3133222
- DOI: 10.1128/MCB.01130-10
Pathways for Holliday junction processing during homologous recombination in Saccharomyces cerevisiae
Abstract
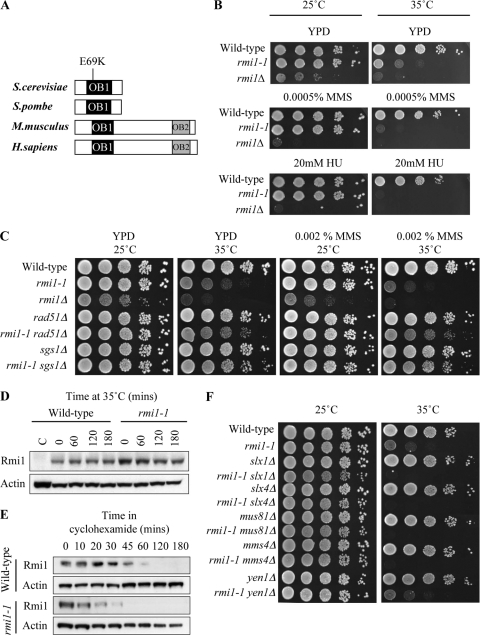
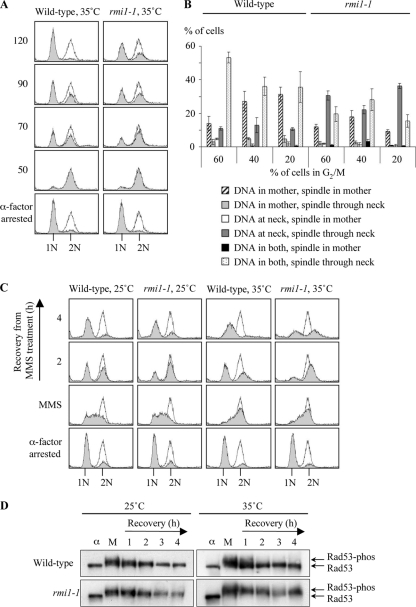
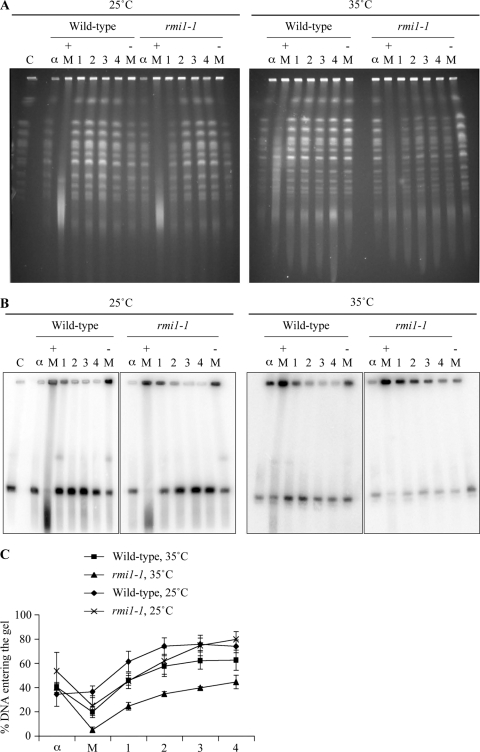
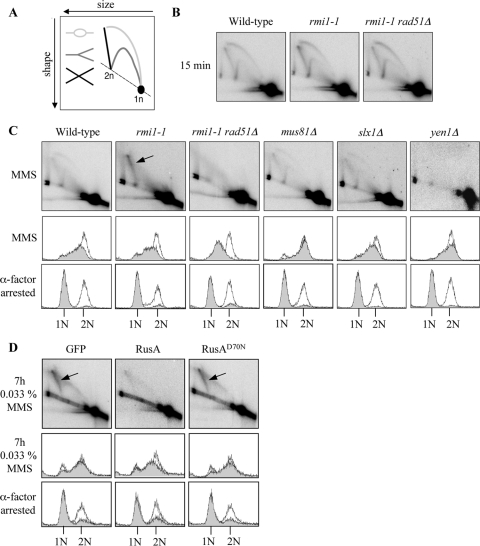
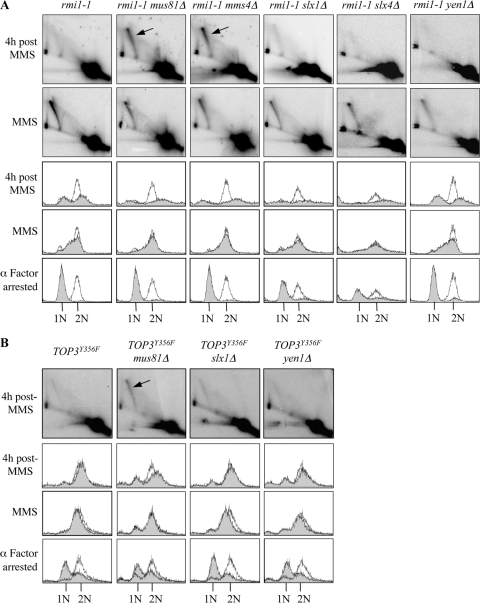
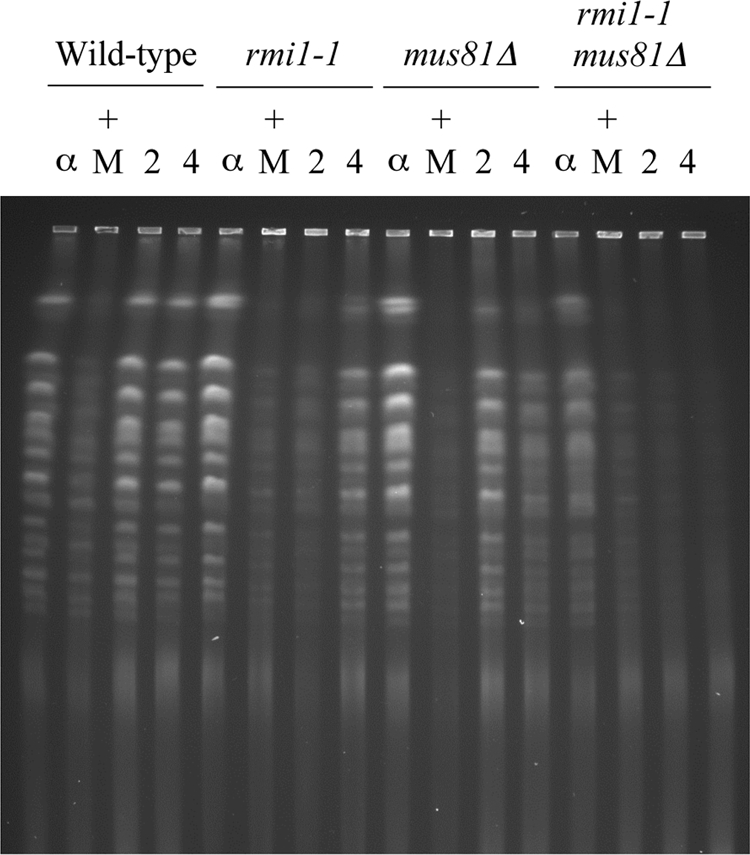
The Saccharomyces cerevisiae Rmi1 protein is a component of the highly conserved Sgs1-Top3-Rmi1 complex. Deletion of SGS1, TOP3, or RMI1 is synthetically lethal when combined with the loss of the Mus81-Mms4 or Slx1-Slx4 endonucleases, which have been implicated in Holliday junction (HJ) resolution. To investigate the causes of this synthetic lethality, we isolated a temperature-sensitive mutant of the RMI1 strain, referred to as the rmi1-1 mutant. At the restrictive temperature, this mutant phenocopies an rmi1Δ strain but behaves like the wild type at the permissive temperature. Following a transient exposure to methyl methanesulfonate, rmi1-1 mutants accumulate unprocessed homologous recombination repair (HRR) intermediates. These intermediates are slowly resolved at the restrictive temperature, revealing a redundant resolution activity when Rmi1 is impaired. This resolution depends on Mus81-Mms4 but not on either Slx1-Slx4 or another HJ resolvase, Yen1. Similar results were also observed when Top3 function was impaired. We propose that the Sgs1-Top3-Rmi1 complex constitutes the main pathway for the processing of HJ-containing HRR intermediates but that Mus81-Mms4 can also resolve these intermediates.
Figures
References
-
- Amberg D., Burke D., Strathern J. 2005. Methods in yeast genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
-
- Ashton T. M., Hickson I. D. 2010. Yeast as a model system to study RecQ helicase function. DNA Repair (Amsterdam) 9:303–314 - PubMed
-
- Bennett R. J., Keck J. L., Wang J. C. 1999. Binding specificity determines polarity of DNA unwinding by the Sgs1 protein of S. cerevisiae. J. Mol. Biol. 289:235–248 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases