

Vacuole-localized berberine bridge enzyme-like proteins are required for a late step of nicotine biosynthesis in tobacco
- PMID: 21343426
- PMCID: PMC3091092
- DOI: 10.1104/pp.110.170878
Vacuole-localized berberine bridge enzyme-like proteins are required for a late step of nicotine biosynthesis in tobacco
Abstract
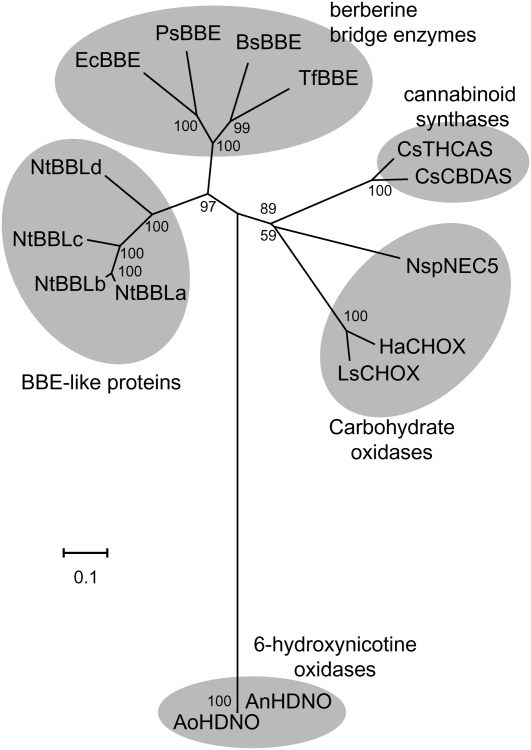
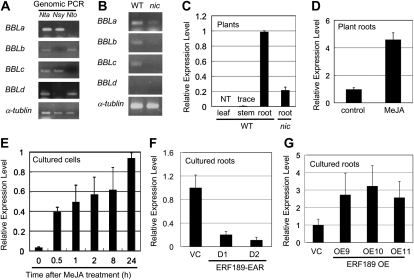
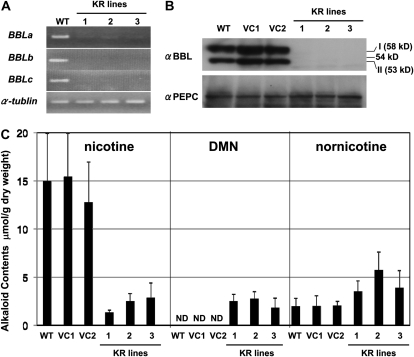
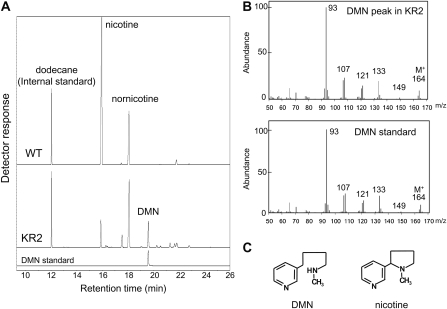
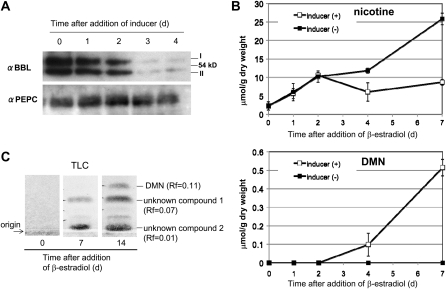
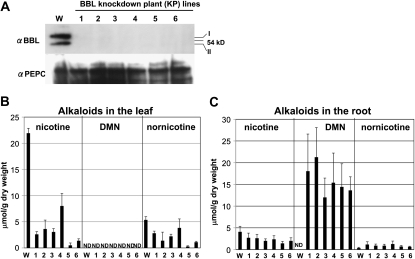
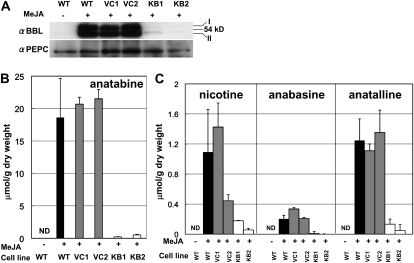
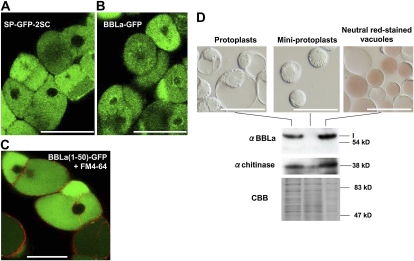
Tobacco (Nicotiana tabacum) plants synthesize nicotine and related pyridine-type alkaloids, such as anatabine, in their roots and accumulate them in their aerial parts as chemical defenses against herbivores. Herbivory-induced jasmonate signaling activates structural genes for nicotine biosynthesis and transport by way of the NICOTINE (NIC) regulatory loci. The biosynthesis of tobacco alkaloids involves the condensation of an unidentified nicotinic acid-derived metabolite with the N-methylpyrrolinium cation or with itself, but the exact enzymatic reactions and enzymes involved remain unclear. Here, we report that jasmonate-inducible tobacco genes encoding flavin-containing oxidases of the berberine bridge enzyme family (BBLs) are expressed in the roots and regulated by the NIC loci. When expression of the BBL genes was suppressed in tobacco hairy roots or in tobacco plants, nicotine production was highly reduced, with a gradual accumulation of a novel nicotine metabolite, dihydromethanicotine. In the jasmonate-elicited cultured tobacco cells, suppression of BBL expression efficiently inhibited the formation of anatabine and other pyridine alkaloids. Subcellular fractionation and localization of green fluorescent protein-tagged BBLs showed that BBLs are localized in the vacuoles. These results indicate that BBLs are involved in a late oxidation step subsequent to the pyridine ring condensation reaction in the biosynthesis of tobacco alkaloids.
Figures

References
-
- Bird DA, Facchini PJ. (2001) Berberine bridge enzyme, a key branch-point enzyme in benzylisoquinoline alkaloid biosynthesis, contains a vacuolar sorting determinant. Planta 213: 888–897 - PubMed
-
- Brandsch R, Hinkkanen AE, Mauch L, Nagursky H, Decker K. (1987) 6-Hydroxy-D-nicotine oxidase of Arthrobacter oxidans: gene structure of the flavoenzyme and its relationship to 6-hydroxy-L-nicotine oxidase. Eur J Biochem 167: 315–320 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
