

Regulation of HIF-1{alpha} activity in adipose tissue by obesity-associated factors: adipogenesis, insulin, and hypoxia
- PMID: 21343542
- PMCID: PMC3093977
- DOI: 10.1152/ajpendo.00626.2010
Regulation of HIF-1{alpha} activity in adipose tissue by obesity-associated factors: adipogenesis, insulin, and hypoxia
Abstract
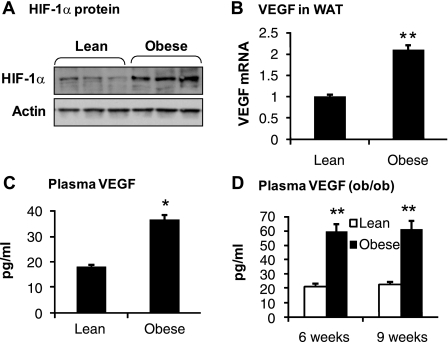
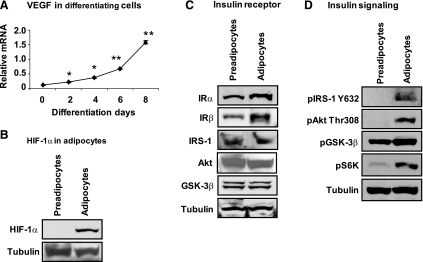
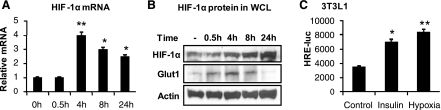
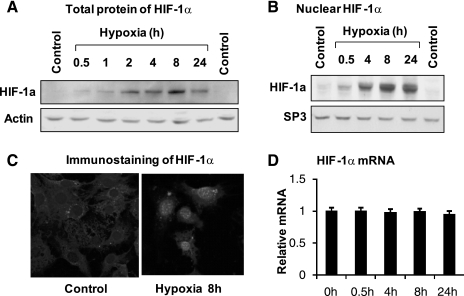
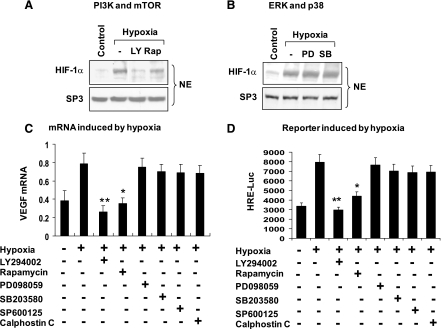
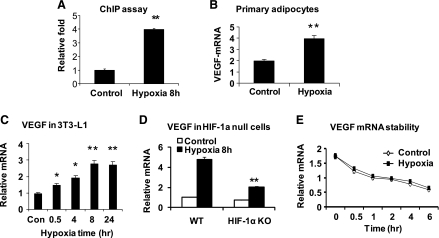
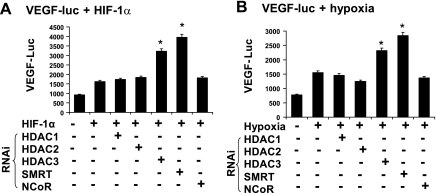
The transcription factor HIF-1α activity is increased in adipose tissue to contribute to chronic inflammation in obesity. However, its upstream and downstream events remain to be characterized in adipose tissue in obesity. We addressed this issue by investigating adipocyte HIF-1α activity in response to obesity-associated factors, such as adipogenesis, insulin, and hypoxia. In adipose tissue, both HIF-1α mRNA and protein were increased by obesity. The underlying mechanism was investigated in 3T3-L1 adipocytes. HIF-1α mRNA and protein were augmented by adipocyte differentiation. In differentiated adipocytes, insulin further enhanced HIF-1α in both levels. Hypoxia enhanced only HIF-1α protein, not mRNA. PI3K and mTOR activities are required for the HIF-1α expression. Function of HIF-1α protein was investigated in the regulation of VEGF gene transcription. ChIP assay shows that HIF-1α binds to the proximal hypoxia response element in the VEGF gene promoter, and its function is inhibited by a corepressor composed of HDAC3 and SMRT. These observations suggest that of the three obesity-associated factors, all of them are able to augment HIF-1α protein levels, but only two (adipogenesis and insulin) are able to enhance HIF-1α mRNA activity. Adipose tissue HIF-1α activity is influenced by multiple signals, including adipogenesis, insulin, and hypoxia in obesity. The transcriptional activity of HIF-1α is inhibited by HDAC3-SMRT corepressor in the VEGF gene promoter.
Figures
References
-
- Arany Z, Foo SY, Ma Y, Ruas JL, Bommi-Reddy A, Girnun G, Cooper M, Laznik D, Chinsomboon J, Rangwala SM, Baek KH, Rosenzweig A, Spiegelman BM. HIF-independent regulation of VEGF and angiogenesis by the transcriptional coactivator PGC-1alpha. Nature 451: 1008–1012, 2008 - PubMed
-
- Bedogni B, Welford SM, Cassarino DS, Nickoloff BJ, Giaccia AJ, Powell MB. The hypoxic microenvironment of the skin contributes to Akt-mediated melanocyte transformation. Cancer Cell 8: 443–454, 2005 - PubMed
-
- Bradbury D, Clarke D, Seedhouse C, Corbett L, Stocks J, Knox A. Vascular endothelial growth factor induction by prostaglandin E2 in human airway smooth muscle cells is mediated by E prostanoid EP2/EP4 receptors and SP-1 transcription factor binding sites. J Biol Chem 280: 29993–30000, 2005 - PubMed
-
- Bråkenhielm E, Cao R, Gao B, Angelin B, Cannon B, Parini P, Cao Y. Angiogenesis inhibitor, TNP-470, prevents diet-induced and genetic obesity in mice. Circ Res 94: 1579–1588, 2004 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous