

The replication rate of human hematopoietic stem cells in vivo
- PMID: 21343613
- PMCID: PMC3099568
- DOI: 10.1182/blood-2010-08-303537
The replication rate of human hematopoietic stem cells in vivo
Abstract
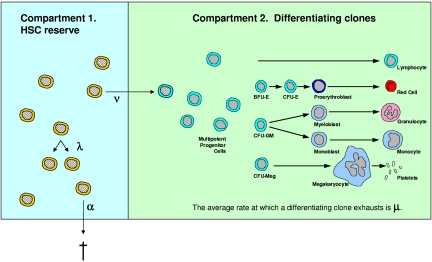
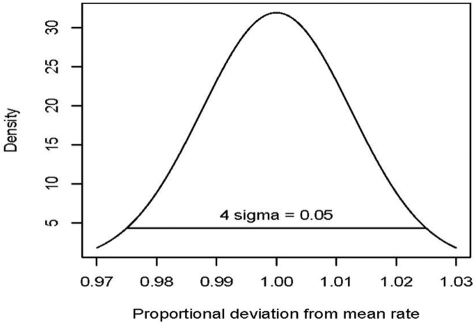
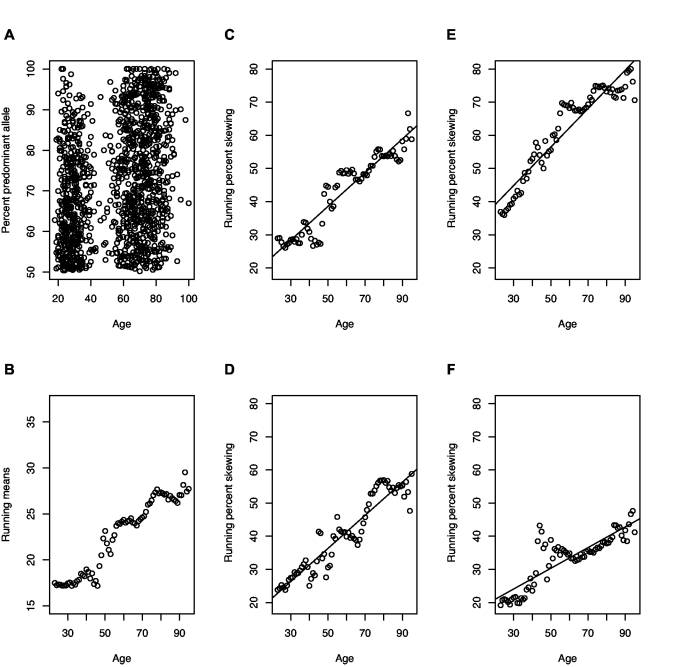
Hematopoietic stem cells (HSCs) replicate (self-renew) to create 2 daughter cells with capabilities equivalent to their parent, as well as differentiate, and thus can both maintain and restore blood cell production. Cell labeling with division-sensitive markers and competitive transplantation studies have been used to estimate the replication rate of murine HSCs in vivo. However, these methods are not feasible in humans and surrogate assays are required. In this report, we analyze the changing ratio with age of maternal/paternal X-chromosome phenotypes in blood cells from females and infer that human HSCs replicate on average once every 40 weeks (range, 25-50 weeks). We then confirm this estimate with 2 independent approaches, use the estimate to simulate human hematopoiesis, and show that the simulations accurately reproduce marrow transplantation data. Our simulations also provide evidence that the number of human HSCs increases from birth until adolescence and then plateaus, and that the ratio of contributing to quiescent HSCs in humans significantly differs from mouse. In addition, they suggest that human marrow failure, such as the marrow failure that occurs after umbilical cord blood transplantation and with aplastic anemia, results from insufficient numbers of early progenitor cells, and not the absence of HSCs.
Figures
References
-
- Schroeder T. Hematopoietic stem cell heterogeneity: subtypes, not unpredictable behavior. Cell Stem Cell. 2010;6(3):203–207. - PubMed
-
- Blank U, Karlsson G, Karlsson S. Signaling pathways governing stem-cell fate. Blood. 2008;111(2):492–503. - PubMed
-
- Raaijmakers MH, Scadden DT. Evolving concepts on the microenvironmental niche for hematopoietic stem cells. Curr Opin Hematol. 2008;15(4):301–306. - PubMed
-
- Guttorp P. Stochastic Modeling of Scientific Data. London, United Kingdom: Chapman and Hall; 1995.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
