

Endocytosis of EphA receptors is essential for the proper development of the retinocollicular topographic map
- PMID: 21343910
- PMCID: PMC3102278
- DOI: 10.1038/emboj.2011.44
Endocytosis of EphA receptors is essential for the proper development of the retinocollicular topographic map
Abstract
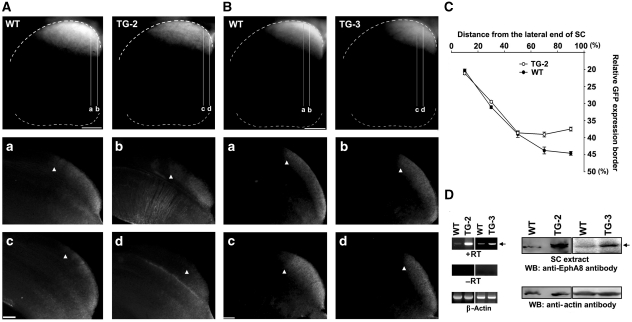
Endocytosis of Eph-ephrin complexes may be an important mechanism for converting cell-cell adhesion to a repulsive interaction. Here, we show that an endocytosis-defective EphA8 mutant forms a complex with EphAs and blocks their endocytosis in cultured cells. Further, we used bacterial artificial chromosome transgenic (Tg) mice to recapitulate the anterior>posterior gradient of EphA in the superior colliculus (SC). In mice expressing the endocytosis-defective EphA8 mutant, the nasal axons were aberrantly shifted to the anterior SC. In contrast, in Tg mice expressing wild-type EphA8, the nasal axons were shifted to the posterior SC, as predicted for the enhanced repellent effect of ephrinA reverse signalling. Importantly, Rac signalling was shown to be essential for EphA-ephrinA internalization and the subsequent nasal axonal repulsion in the SC. These results indicate that endocytosis of the Eph-ephrin complex is a key mechanism by which axonal repulsion is generated for proper guidance and topographic mapping.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures








Comment in
-
D-Eph-ective endocytosis disrupts topographic mapping.EMBO J. 2011 Apr 20;30(8):1422-4. doi: 10.1038/emboj.2011.87. EMBO J. 2011. PMID: 21505521 Free PMC article.
Similar articles
-
Eph and ephrin signaling in the formation of topographic maps.Semin Cell Dev Biol. 2012 Feb;23(1):7-15. doi: 10.1016/j.semcdb.2011.10.026. Epub 2011 Oct 24. Semin Cell Dev Biol. 2012. PMID: 22044886 Free PMC article. Review.
-
D-Eph-ective endocytosis disrupts topographic mapping.EMBO J. 2011 Apr 20;30(8):1422-4. doi: 10.1038/emboj.2011.87. EMBO J. 2011. PMID: 21505521 Free PMC article.
-
Protein Tyrosine Phosphatase Receptor Type J (PTPRJ) Regulates Retinal Axonal Projections by Inhibiting Eph and Abl Kinases in Mice.J Neurosci. 2018 Sep 26;38(39):8345-8363. doi: 10.1523/JNEUROSCI.0128-18.2018. Epub 2018 Aug 6. J Neurosci. 2018. PMID: 30082414 Free PMC article.
-
The L1 cell adhesion molecule is essential for topographic mapping of retinal axons.J Neurosci. 2003 Jan 15;23(2):530-8. doi: 10.1523/JNEUROSCI.23-02-00530.2003. J Neurosci. 2003. PMID: 12533613 Free PMC article.
-
Axonal ephrinA/EphA interactions, and the emergence of order in topographic projections.Semin Cell Dev Biol. 2012 Feb;23(1):1-6. doi: 10.1016/j.semcdb.2011.10.015. Epub 2011 Oct 21. Semin Cell Dev Biol. 2012. PMID: 22040913 Review.
Cited by
-
Over-Expression of Ephrin-A5 in Mice Results in Decreasing the Size of Progenitor Pool through Inducing Apoptosis.Mol Cells. 2016 Feb;39(2):136-40. doi: 10.14348/molcells.2016.2245. Epub 2015 Dec 15. Mol Cells. 2016. PMID: 26674965 Free PMC article.
-
Eph/ephrin signaling in epidermal differentiation and disease.Semin Cell Dev Biol. 2012 Feb;23(1):92-101. doi: 10.1016/j.semcdb.2011.10.017. Epub 2011 Oct 21. Semin Cell Dev Biol. 2012. PMID: 22040910 Free PMC article. Review.
-
EphA3 expressed in the chicken tectum stimulates nasal retinal ganglion cell axon growth and is required for retinotectal topographic map formation.PLoS One. 2012;7(6):e38566. doi: 10.1371/journal.pone.0038566. Epub 2012 Jun 7. PLoS One. 2012. PMID: 22685584 Free PMC article.
-
Eph and ephrin signaling in the formation of topographic maps.Semin Cell Dev Biol. 2012 Feb;23(1):7-15. doi: 10.1016/j.semcdb.2011.10.026. Epub 2011 Oct 24. Semin Cell Dev Biol. 2012. PMID: 22044886 Free PMC article. Review.
-
Ephrin-A3 is required for tonotopic map precision and auditory functions in the mouse auditory brainstem.J Comp Neurol. 2021 Nov;529(16):3633-3654. doi: 10.1002/cne.25213. Epub 2021 Jul 16. J Comp Neurol. 2021. PMID: 34235739 Free PMC article.
References
-
- Batlle E, Bacani J, Begthel H, Jonkheer S, Gregorieff A, van de Born M, Malats N, Sancho E, Boon E, Pawson T, Gallinger S, Pals S, Clevers H (2005) EphB receptor activity suppresses colorectal cancer progression. Nature 435: 1126–1130 - PubMed
-
- Bruckner K, Klein R (1998) Signaling by Eph receptors and their ephrin ligands. Curr Opin Neurobiol 8: 375–382 - PubMed
-
- Choi S, Park S (1999) Phosphorylation at Tyr-838 in the kinase domain of EphA8 modulates Fyn binding to the Tyr-615 site by enhancing tyrosine kinase activity. Oncogene 18: 5413–5422 - PubMed
-
- Cowan CA, Henkemeyer M (2001) The SH2/SH3 adaptor Grb4 transduces B-ephrin reverse signals. Nature 413: 174–179 - PubMed
-
- Egea J, Klein R (2007) Bidirectional Eph-ephrin signaling during axon guidance. Trends Cell Biol 17: 230–238 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
