

The actin cytoskeleton has an active role in the electrotransfer of plasmid DNA in mammalian cells
- PMID: 21343915
- PMCID: PMC3098633
- DOI: 10.1038/mt.2010.303
The actin cytoskeleton has an active role in the electrotransfer of plasmid DNA in mammalian cells
Abstract
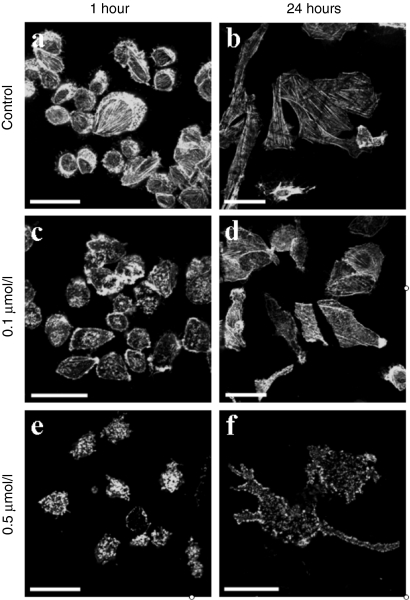
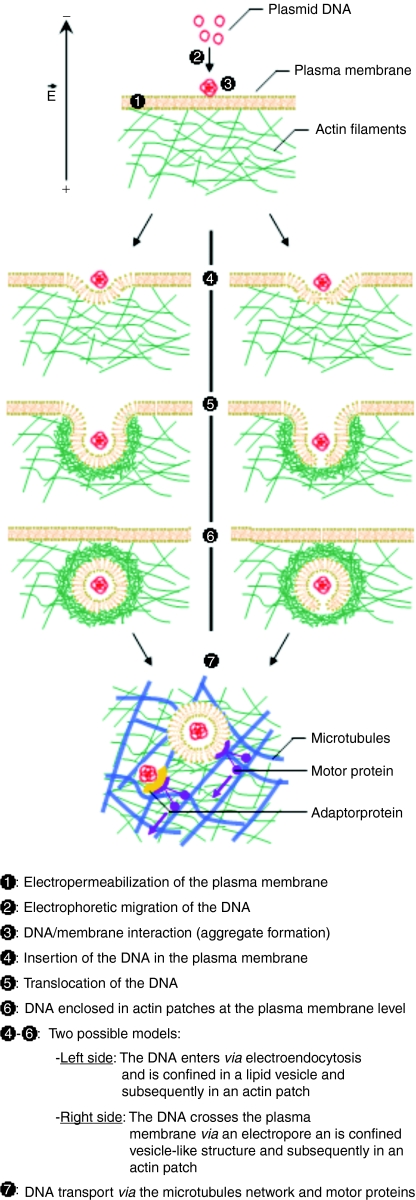
Electrotransfer of molecules is a well established technique which finds extensive use for gene transfer and holds great promise for anticancer treatment. Despite its widespread application, the mechanisms governing the entry of DNA into the cell and its intracellular trafficking are not yet known. The aim of this study is to unravel the role of the actin cytoskeleton during gene electrotransfer in cells. We performed single-cell level approaches to observe the organization of the actin cytoskeleton in Chinese hamster ovary (CHO) cells. In addition, we performed experiments at the multiple-cell level to evaluate the efficiency of DNA transfer after alteration of the actin cytoskeleton using the drug latrunculin B. Actin patches colocalizing with the DNA at the plasma membrane were observed with additional characteristics similar to those of the DNA aggregates in terms of time, number, and size. The disruption of the microfilaments reduces the DNA accumulation at the plasma membrane and the gene expression. This is the first direct experimental evidence of the participation of the actin cytoskeleton in DNA electrotransfer.
Figures





References
-
- Young LS, Searle PF, Onion D., and, Mautner V. Viral gene therapy strategies: from basic science to clinical application. J Pathol. 2006;208:299–318. - PubMed
-
- Karmali PP., and, Chaudhuri A. Cationic liposomes as non-viral carriers of gene medicines: resolved issues, open questions, and future promises. Med Res Rev. 2007;27:696–722. - PubMed
-
- Newman CM., and, Bettinger T. Gene therapy progress and prospects: ultrasound for gene transfer. Gene Ther. 2007;14:465–475. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
