

Review
doi: 10.1016/j.virusres.2011.02.011.
Epub 2011 Feb 21.
Molecular virology of hepatitis E virus
Affiliations
- PMID: 21345356
- PMCID: PMC3130092
- DOI: 10.1016/j.virusres.2011.02.011
Item in Clipboard
Review
Molecular virology of hepatitis E virus
Virus Res.
2011 Oct.
Abstract
This review details the molecular virology of the hepatitis E virus (HEV). While replicons and in vitro infection systems have recently become available, a lot of information on HEV has been generated through comparisons with better-studied positive-strand RNA viruses and through subgenomic expression of viral open reading frames. These models are now being verified with replicon and infection systems. We provide here the current knowledge on the HEV genome and its constituent proteins--ORF1, ORF2 and ORF3. Based on the available information, we also modify the existing model of the HEV life cycle.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures

The hepatitis E virus genome. The ∼7.2 kb positive sense RNA genome of HEV has a 7-Me-G cap at its 5′ end and a poly A tail at its 3′ end. There are short stretches of untranslated regions at the 5′ and 3′ ends that fold into stem-loop structures (shown in blue). The three open reading frames are shown. ORF1 encodes a nonstructural polyprotein and contains a 58-nucleotide stretch near its 5′ end that folds into a stem-loop structure (shown in green). The ORF2 and ORF3 proteins are translated from a 2.2 kb subgenomic RNA generated during viral replication. The boxed region on the upper right shows the sequence alignment of the junction region between orf1 and orf3 in HEV isolates representative of genotypes 1–4. The nucleotide positions are shown with respect to HEV genotype 1 (Sar55). Dots indicate identity and dashes represent deletions. The orf1 stop codon is shown in red. There is a single nucleotide insertion (T, indicated with filled triangle) between positions 5116 and 5117 in HEV genotype 4. The four initiation codons within this junction region are shown in yellow boxes, and are at positions 5104, 5113, 5131 and 5145. This region has been predicted to fold into a double stem-loop structure shown in the boxed region on the left. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)

The ORF1 protein. The orf1 gene encodes a nonstructural polyprotein with four predicted functional domains, designated as methyltransferase (MeT), papain-like cysteine protease (PCP), helicase (Hel) and RNA-dependent RNA polymerase (RdRp). Besides these regions, two other regions designated X (macro domain) and Y share significant homology with nonstructural proteins of other positive-strand RNA viruses. A proline-rich region (V) upstream of the macro domain may act as a flexible hinge. Numbers indicate the predicted boundaries of the different regions; these are based on the genotype 1 Burmese isolate.

The ORF2 protein. The orf2 gene encodes the HEV capsid protein. The full-length protein is 660 amino acids long. In animal cells, pORF2 is glycosylated and three N-linked glycosylation sites have been mapped to the indicated asparagine (Asn) residues. A truncated protein of 56 kDa (amino acids 112–607) can self-assemble in insect cells to form virus-like particles (VLPs), which possess the same dominant antigenic epitopes as the virion. Various shorter constructs have been expressed in E. coli. Of these, p239 forms particles via dimeric interactions while E2 and E2a form hexameric surface protrusions. The E2s is proposed as the dimerization domain. Two of these proteins (yellow highlight) have undergone clinical testing in humans as HEV vaccine candidates. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
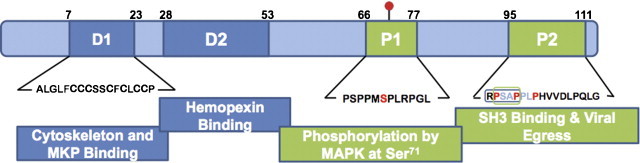
The ORF3 protein. The orf3 gene encodes a 114-amino acid multi-functional protein, which has two hydrophobic domains (D1 and D2) and two proline rich domains (P1 and P2). In mammalian cells, the ORF3 protein is phosphorylated at Serine 71 (red dot) by cellular MAPK. The proline-rich P2 domain contains a PXXP motif (RPSAP, in box) that is implicated in binding to SH3 domains in cellular proteins. An intact PSAP motif is also required for viral egress. Regions of the ORF3 protein experimentally shown to be involved in various interactions are indicated. Numbers represent the proposed boundaries of the various domains. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)

Proposed replication cycle of hepatitis E virus. The viral particles are concentrated on the surface of target cells through heparin sulfate proteoglycans acting as attachment factors (red wavy lines) (step 1) and subsequently bind a specific yet uncharacterized receptor (step 2), following which the particles are internalized (step 3). The virus then uncoats (step 4) to release genomic RNA (red noodles) that is translated in the cytoplasm into nonstructural proteins (step 5). These nonstructural proteins include the RNA dependent RNA polymerase that replicates the positive sense genomic RNA into negative sense transcripts (purple noodles) (step 6); the latter then act as templates for the synthesis of a 2.2 kb subgenomic RNA (step 7a) as well as full-length positive sense transcripts (step 7b). The positive sense subgenomic RNA is translated into ORF2 (blue) and ORF3 (crimson) proteins (step 8). The ORF2 protein packages the genomic RNA to assemble new virions (step 9) while the ORF3 protein may optimize the host cell environment for viral replication. The ORF3 protein is also associated with endomembranes (step 10a) or plasma membranes (step 10b) and may aid in viral egress. Recent studies suggest that mature virions are associated with the ORF3 protein and lipids (step 11), which are subsequently removed through a process that is not understood at present, to resume a fresh infection cycle. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
References
-
- Agrawal S., Gupta D., Panda S.K. The 3′ end of hepatitis E virus (HEV) genome binds specifically to the viral RNA-dependent RNA polymerase (RdRp) Virology. 2001;282:87–101. - PubMed
-
- Aguiar R.C., Takeyama K., He C., Kreinbrink K., Shipp M.A. B-aggressive lymphoma family proteins have unique domains that modulate transcription and exhibit poly(ADP-ribose) polymerase activity. J. Biol. Chem. 2005;280:33756–33765. - PubMed
-
- Ansari I.H., Nanda S.K., Durgapal H., Agrawal S., Mohanty S.K., Gupta D., Jameel S., Panda S.K. Cloning, sequencing, and expression of the hepatitis E virus (HEV) nonstructural open reading frame 1 (ORF1) J. Med. Virol. 2000;60:275–283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
