

Prefrontal cortical network connections: key site of vulnerability in stress and schizophrenia
- PMID: 21345366
- PMCID: PMC3115784
- DOI: 10.1016/j.ijdevneu.2011.02.006
Prefrontal cortical network connections: key site of vulnerability in stress and schizophrenia
Abstract
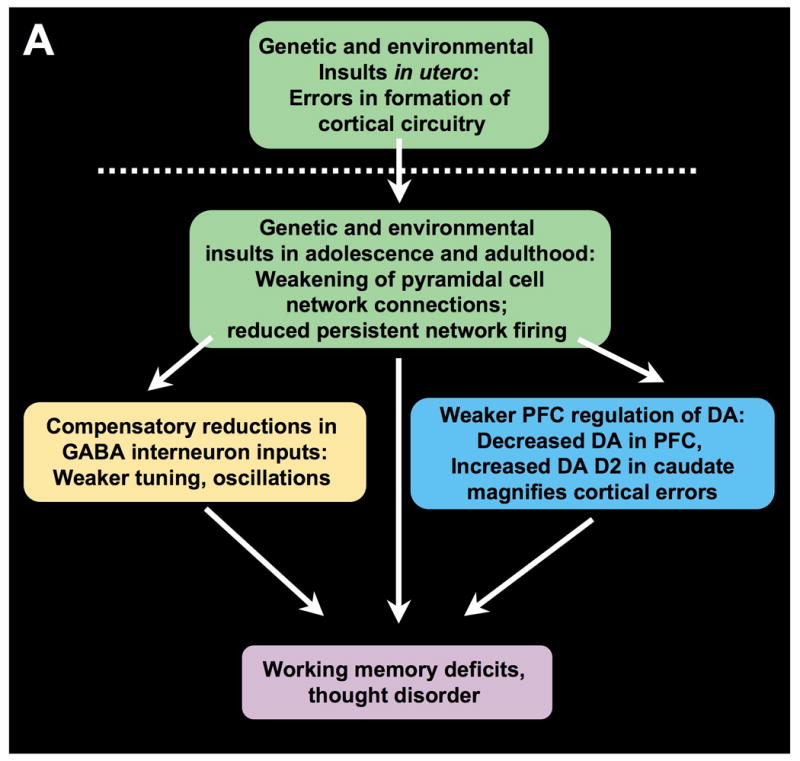
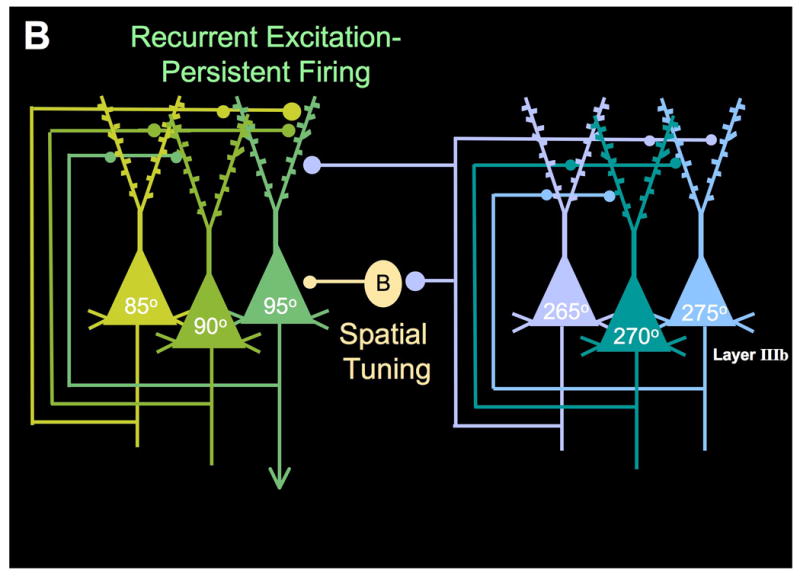
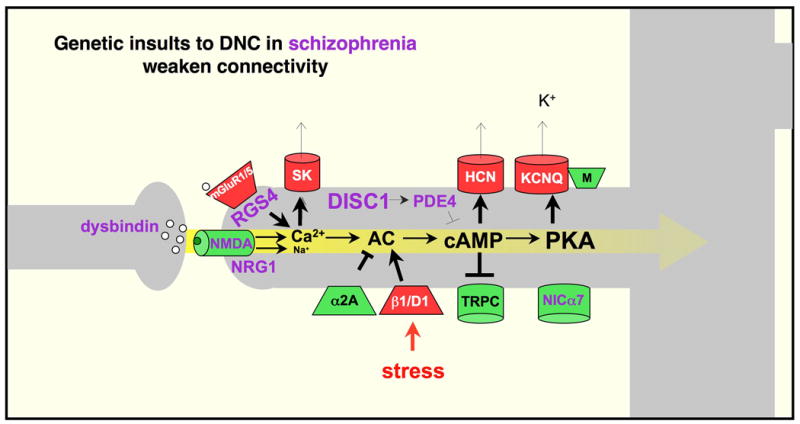
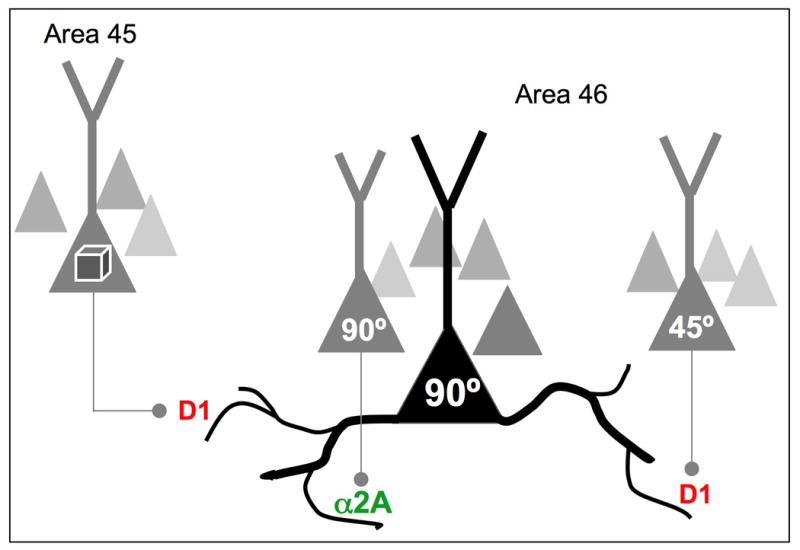
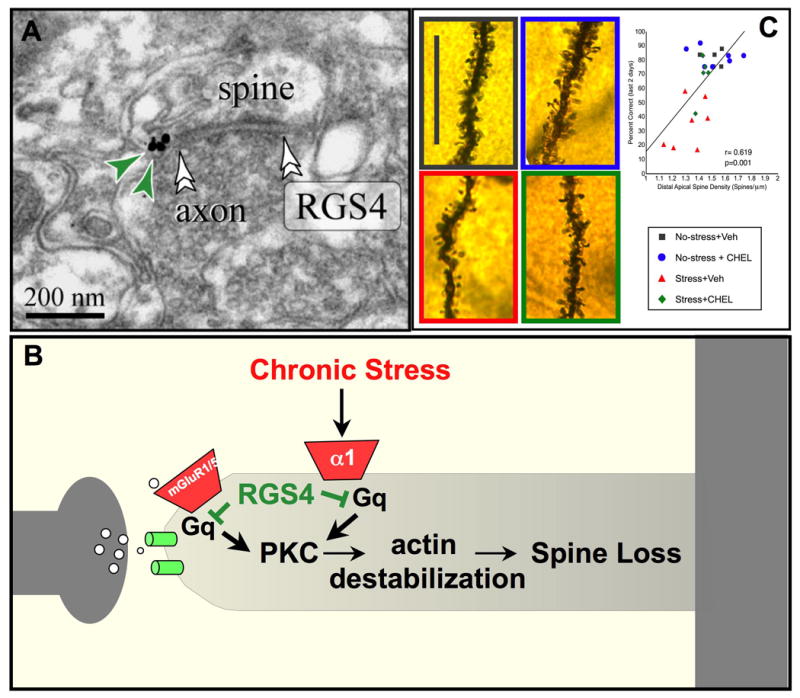
The symptoms of schizophrenia involve profound dysfunction of the prefrontal cortex (PFC). PFC networks create our "mental sketch pad", and PFC dysfunction contributes to symptoms such as cognitive deficits, thought disorder, delusions and hallucinations. Neuropathological studies of schizophrenia have shown marked loss of dendritic spines in deep layer III, the sublayer where PFC microcircuits reside. The microcircuits consist of recurrent excitatory pyramidal cell networks that interconnect on spines, and excite each other via NMDA receptor signaling. The pyramidal cell persistent firing is sculpted by lateral inhibition from GABAergic basket and chandelier cells, thus creating tuned, persistent firing needed for accurate representational knowledge (i.e., working memory). The strength of pyramidal cell network connections is markedly and flexibly altered by intracellular signaling pathways in dendritic spines, a process called dynamic network connectivity (DNC). DNC proteins such as HCN channels are concentrated on dendritic spines in deep layer III. Under optimal conditions, network inputs to pyramidal cells are strengthened by noradrenergic alpha-2A inhibition of cAMP-HCN channel signaling, and sculpted by dopamine D1-cAMP-HCN channel weakening of inappropriate inputs. However, with stress exposure, high levels of cAMP-HCN channel signaling produces a collapse in network firing. With chronic stress exposure, spines reduce in size and are lost, and this process involves increased PKC signaling. Importantly, molecules that normally strengthen PFC networks connections and/or reverse the stress response, are often genetically altered in schizophrenia. As exposure to stress is a key factor in the precipitation of schizophrenic symptoms, these dysregulated signaling pathways in deep layer III may interact with already vulnerable circuitry to cause spine loss and the descent into illness.
Copyright © 2011 ISDN. Published by Elsevier Ltd. All rights reserved.
Figures
References
-
- Akil M, Pierri JN, Whitehead RE, Edgar CL, Mohila C, Sampson AR, Lewis DA. Lamina-specific alterations in the dopamine innervation of the prefrontal cortex in schizophrenic subjects. Am J Psychiatry. 1999;156:1580–1589. - PubMed
-
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Ann Rev Neurosci. 1986;9:357–381. - PubMed
-
- Arnsten AFT, Goldman-Rakic PS. Stress impairs prefrontal cortex cognitive function in monkeys: role of dopamine. Soc Neurosci Abstr. 1990;16:164.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
