

Fat cells reactivate quiescent neuroblasts via TOR and glial insulin relays in Drosophila
- PMID: 21346761
- PMCID: PMC3146047
- DOI: 10.1038/nature09867
Fat cells reactivate quiescent neuroblasts via TOR and glial insulin relays in Drosophila
Abstract
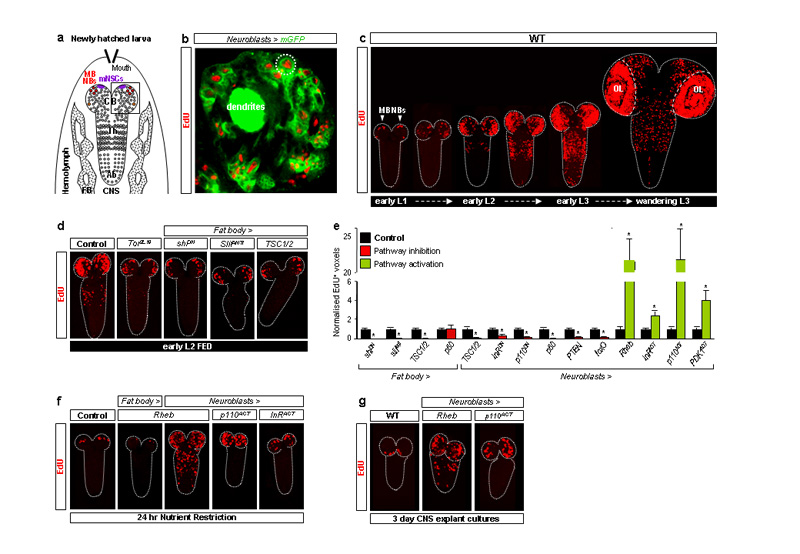
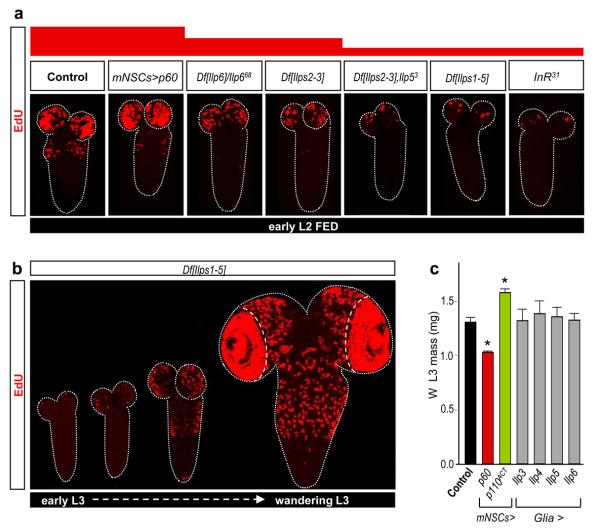
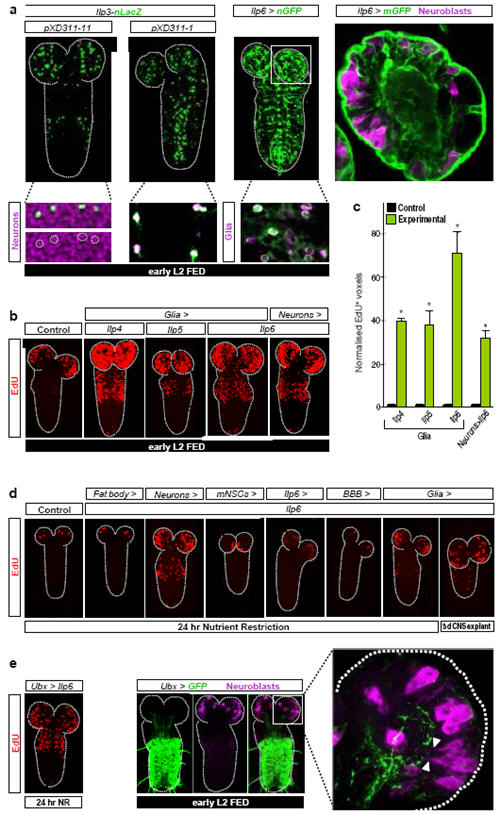
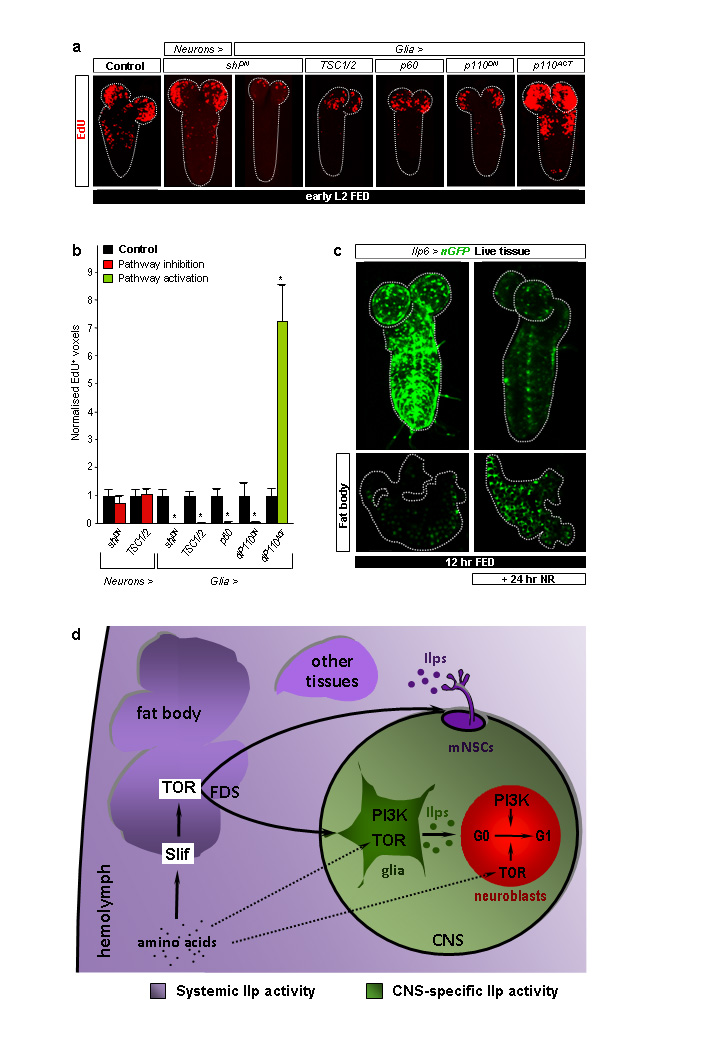
Many stem, progenitor and cancer cells undergo periods of mitotic quiescence from which they can be reactivated. The signals triggering entry into and exit from this reversible dormant state are not well understood. In the developing Drosophila central nervous system, multipotent self-renewing progenitors called neuroblasts undergo quiescence in a stereotypical spatiotemporal pattern. Entry into quiescence is regulated by Hox proteins and an internal neuroblast timer. Exit from quiescence (reactivation) is subject to a nutritional checkpoint requiring dietary amino acids. Organ co-cultures also implicate an unidentified signal from an adipose/hepatic-like tissue called the fat body. Here we provide in vivo evidence that Slimfast amino-acid sensing and Target of rapamycin (TOR) signalling activate a fat-body-derived signal (FDS) required for neuroblast reactivation. Downstream of this signal, Insulin-like receptor signalling and the Phosphatidylinositol 3-kinase (PI3K)/TOR network are required in neuroblasts for exit from quiescence. We demonstrate that nutritionally regulated glial cells provide the source of Insulin-like peptides (ILPs) relevant for timely neuroblast reactivation but not for overall larval growth. Conversely, ILPs secreted into the haemolymph by median neurosecretory cells systemically control organismal size but do not reactivate neuroblasts. Drosophila thus contains two segregated ILP pools, one regulating proliferation within the central nervous system and the other controlling tissue growth systemically. Our findings support a model in which amino acids trigger the cell cycle re-entry of neural progenitors via a fat-body-glia-neuroblasts relay. This mechanism indicates that dietary nutrients and remote organs, as well as local niches, are key regulators of transitions in stem-cell behaviour.
Figures
Comment in
-
Food for thought: neural stem cells on a diet.Cell Stem Cell. 2011 Apr 8;8(4):352-4. doi: 10.1016/j.stem.2011.03.004. Cell Stem Cell. 2011. PMID: 21474096
References
METHODS REFERENCES
References
-
- Dhawan J, Rando TA. Stem cells in postnatal myogenesis: molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends Cell Biol. 2005;15:666–73. - PubMed
-
- Coller HA. What's taking so long? S-phase entry from quiescence versus proliferation. Nat Rev Mol Cell Biol. 2007;8:667–70. - PubMed
-
- Yanagida M. Cellular quiescence: are controlling genes conserved? Trends Cell Biol. 2009;19:705–715. - PubMed
-
- Chen E, Finkel T. Preview. The Tortoise, the hare, and the FoxO. Cell Stem Cell. 2009;5:451–2. - PubMed
-
- Sanchez-Garcia I, Vicente-Duenas C, Cobaleda C. The theoretical basis of cancer-stem-cell-based therapeutics of cancer: can it be put into practice? Bioessays. 2007;29:1269–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
