

Epstein-Barr virus nuclear antigen 3C facilitates G1-S transition by stabilizing and enhancing the function of cyclin D1
- PMID: 21347341
- PMCID: PMC3037348
- DOI: 10.1371/journal.ppat.1001275
Epstein-Barr virus nuclear antigen 3C facilitates G1-S transition by stabilizing and enhancing the function of cyclin D1
Abstract
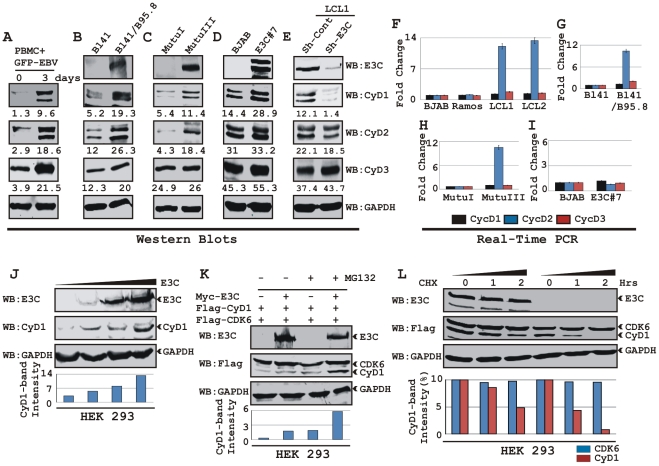
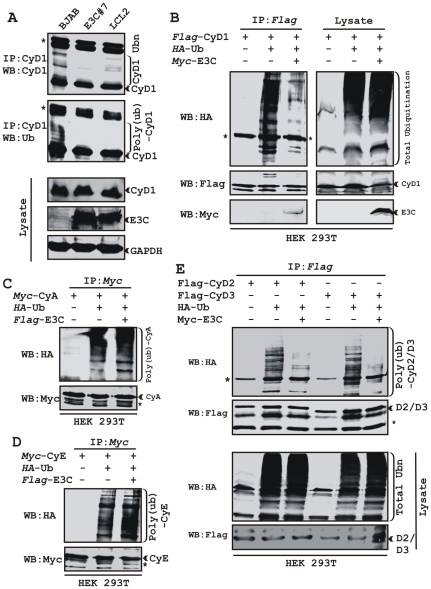
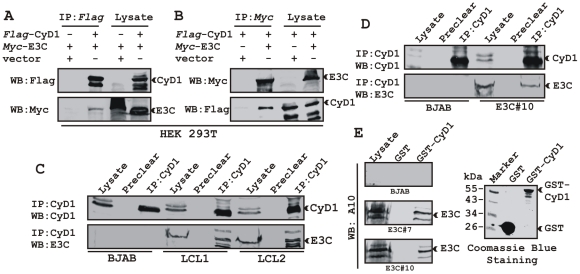
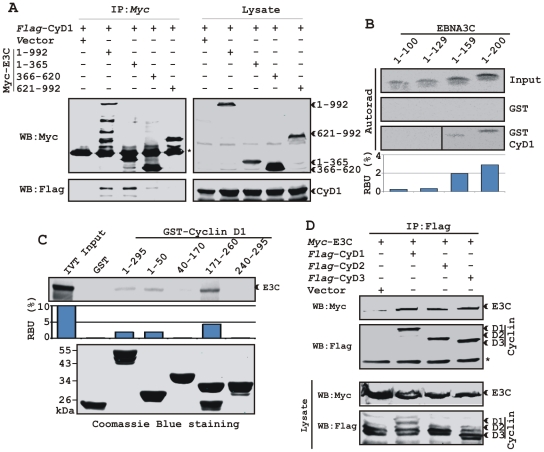
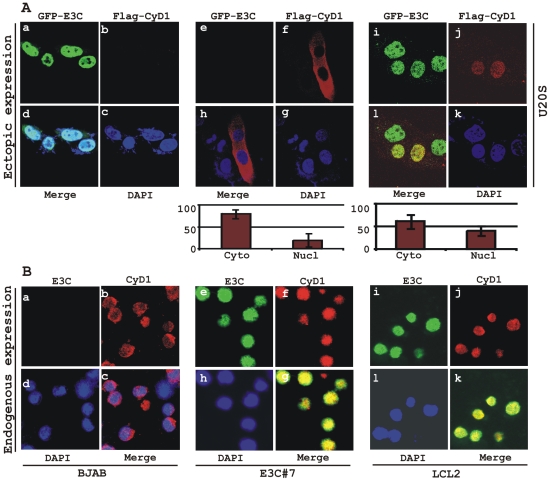
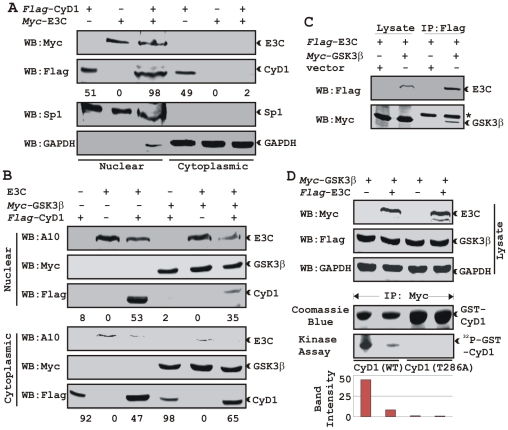
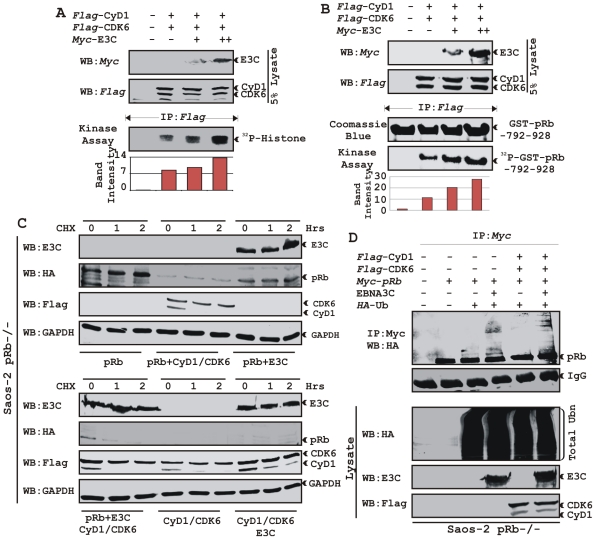
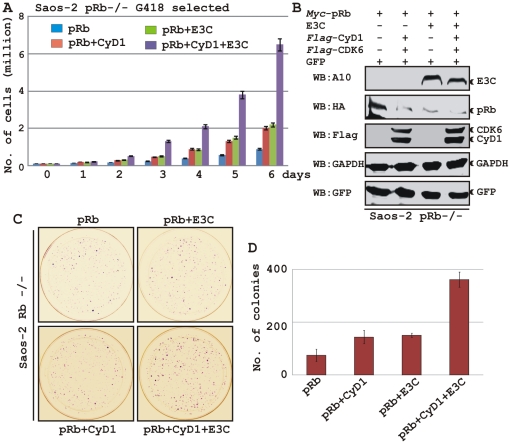
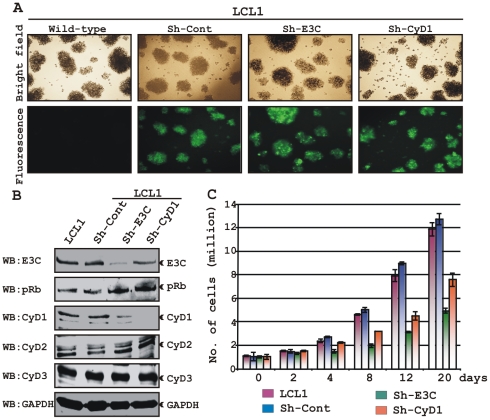
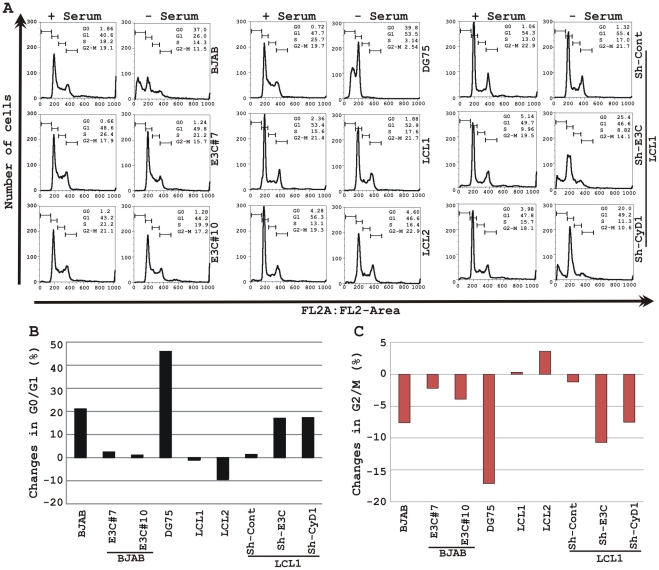
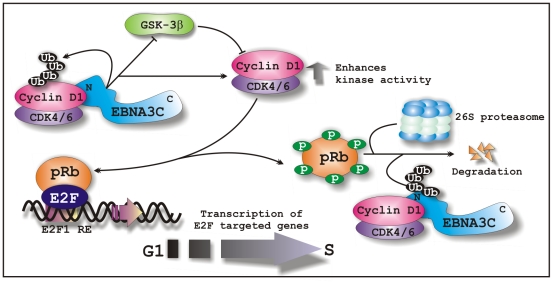
EBNA3C, one of the Epstein-Barr virus (EBV)-encoded latent antigens, is essential for primary B-cell transformation. Cyclin D1, a key regulator of G1 to S phase progression, is tightly associated and aberrantly expressed in numerous human cancers. Previously, EBNA3C was shown to bind to Cyclin D1 in vitro along with Cyclin A and Cyclin E. In the present study, we provide evidence which demonstrates that EBNA3C forms a complex with Cyclin D1 in human cells. Detailed mapping experiments show that a small N-terminal region which lies between amino acids 130-160 of EBNA3C binds to two different sites of Cyclin D1- the N-terminal pRb binding domain (residues 1-50), and C-terminal domain (residues 171-240), known to regulate Cyclin D1 stability. Cyclin D1 is short-lived and ubiquitin-mediated proteasomal degradation has been targeted as a means of therapeutic intervention. Here, we show that EBNA3C stabilizes Cyclin D1 through inhibition of its poly-ubiquitination, and also increases its nuclear localization by blocking GSK3β activity. We further show that EBNA3C enhances the kinase activity of Cyclin D1/CDK6 which enables subsequent ubiquitination and degradation of pRb. EBNA3C together with Cyclin D1-CDK6 complex also efficiently nullifies the inhibitory effect of pRb on cell growth. Moreover, an sh-RNA based strategy for knock-down of both cyclin D1 and EBNA3C genes in EBV transformed lymphoblastoid cell lines (LCLs) shows a significant reduction in cell-growth. Based on these results, we propose that EBNA3C can stabilize as well as enhance the functional activity of Cyclin D1 thereby facilitating the G1-S transition in EBV transformed lymphoblastoid cell lines.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Robertson ES. Norwich, United Kingdom: Caister Academic Press; 2010. Epstein-Barr virus: latency and transformation.
-
- Kieff E, Rickinson AB. Epstein-Barr virus and its replication. In: Knipe D, Howley P, editors. Fields virology, 4th ed. Philadelphia: Lippincott Williams & Wilkins; 2002. pp. 2511–2573.
-
- O'Nions J, Allday MJ. Deregulation of the cell cycle by the Epstein-Barr virus. Adv Cancer Res. 2004;92:119–186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
