

Characterization of the structure and DNA complexity of mung bean mitochondrial nucleoids
- PMID: 21347700
- PMCID: PMC3932694
- DOI: 10.1007/s10059-011-0036-4
Characterization of the structure and DNA complexity of mung bean mitochondrial nucleoids
Abstract
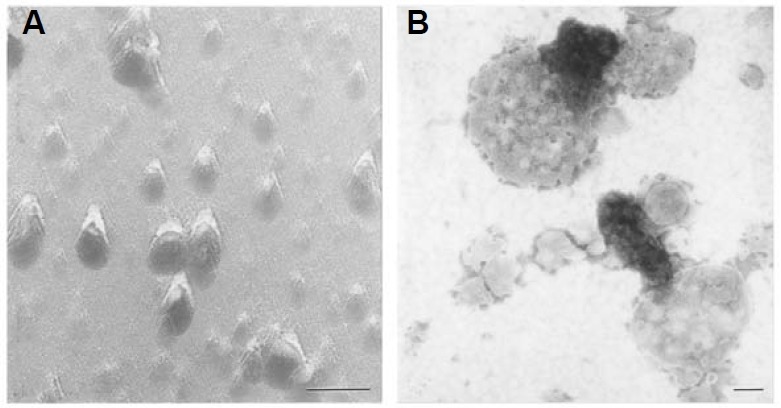
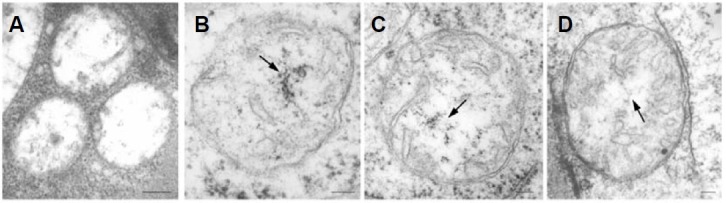
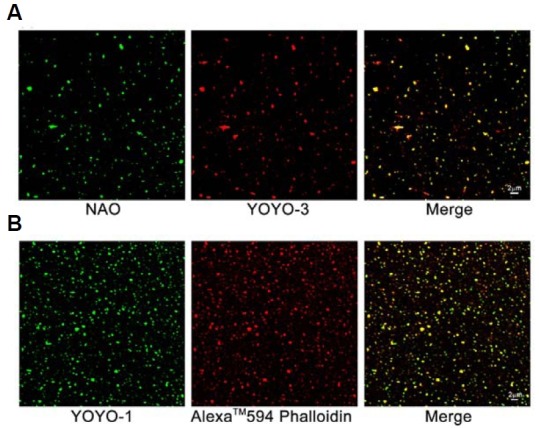
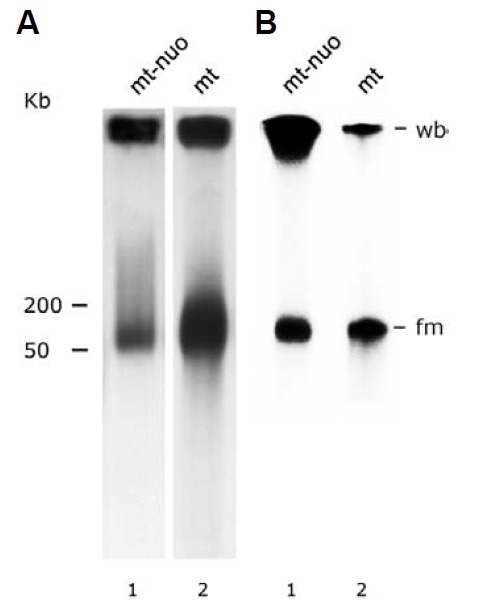
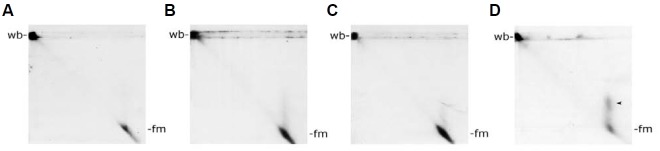
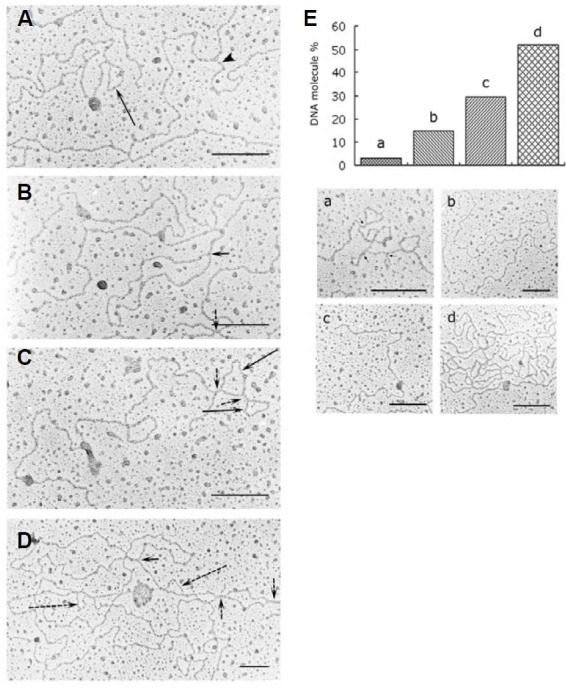
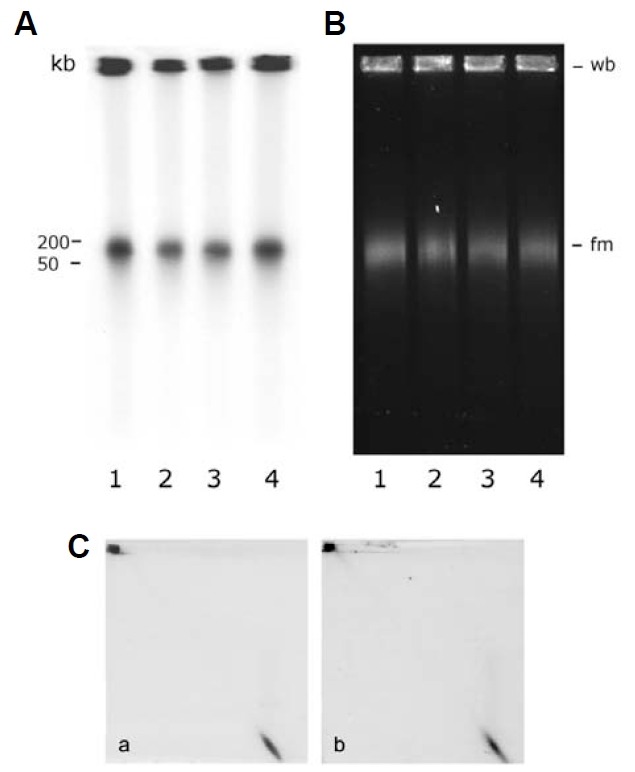
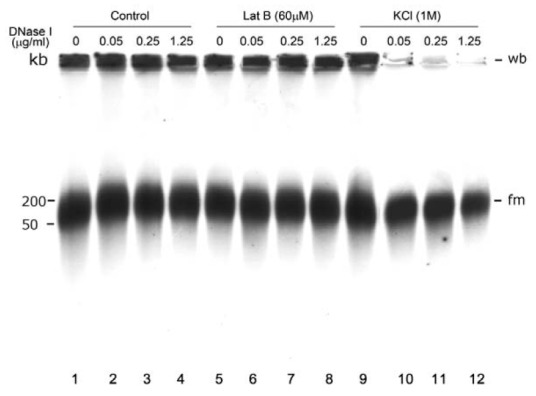
Electron microscopic images of mitochondrial nucleoids isolated from mung bean seedlings revealed a relatively homogeneous population of particles, each consisting of a chromatin-like structure associated with a membrane component. Association of F-actin with mitochondrial nucleoids was also observed. The mitochondrial nucleoid structure identified in situ showed heterogeneous genomic organization. After pulsed-field gel electrophoresis (PFGE), a large proportion of the mitochondrial nucleoid DNA remained in the well, whereas the rest migrated as a 50-200 kb smear zone. This PFGE migration pattern was not affected by high salt, topoisomerase I or latrunculin B treatments; however, the mobility of a fraction of the fast-moving DNA decreased conspicuously following an in-gel ethidium-enhanced UV-irradiation treatment, suggesting that molecules with intricately compact structures were present in the 50-200 kb region. Approximately 70% of the mitochondrial nucleoid DNA molecules examined via electron microscopy were open circles, supercoils, complex forms, and linear molecules with interspersed sigma-shaped structures and/or loops. Increased sensitivity of mtDNA to DNase I was found after mitochondrial nucleoids were pretreated with high salt. This result indicates that some loosely bound or peripheral DNA binding proteins protected the mtDNA from DNase I degradation.
Figures
References
-
- Andre C.P., Walbot V. Pulsed-field gel mapping of maize mitochondrial chromosomes. Mol. Gen. Genet. (1995);247:255–263. - PubMed
-
- Backert S., Börner T. Phage T-4-like intermediates of DNA replication and recombination in the mitochondria of the higher plant Chenopodium album (L.). Curr. Genet. (2000);37:304–314. - PubMed
-
- Backert S., Lurz R., Börner T. Electron micro-scopic investigation of mitochondrial DNA from Chenopodium album (L.). Curr. Genet. (1996);29:427–436. - PubMed
-
- Backert S., Lurz R., Oyarzabal O.A., Borner T. High content, size and distribution of single-stranded DNA in the mitochondria of Chenopodium album (L.). Plant Mol. Biol. (1997);33:1037–1050. - PubMed
-
- Bendich A.J. Structural analysis of mitochondrial DNA molecules from fungi and plants using moving pictures and pulsed-field gel electrophoresis. J. Mol. Biol. (1996);255:564–588. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
