

Stochastic exit from mitosis in budding yeast: model predictions and experimental observations
- PMID: 21350333
- PMCID: PMC3100879
- DOI: 10.4161/cc.10.6.14966
Stochastic exit from mitosis in budding yeast: model predictions and experimental observations
Abstract
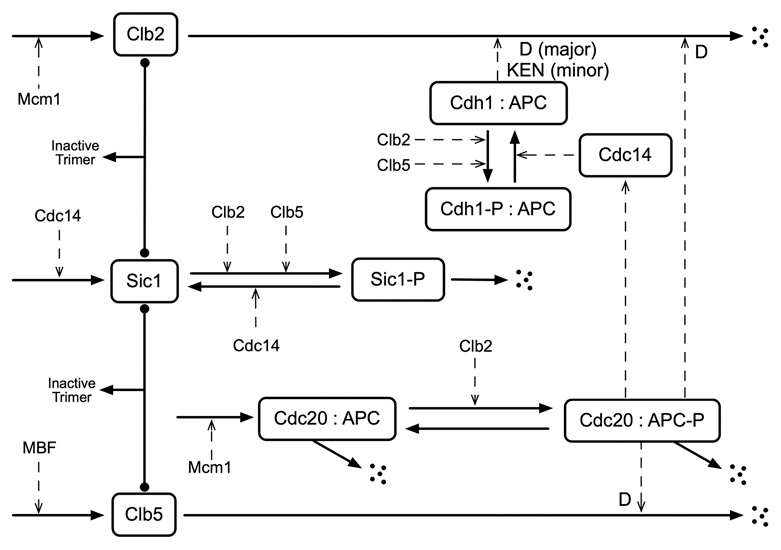
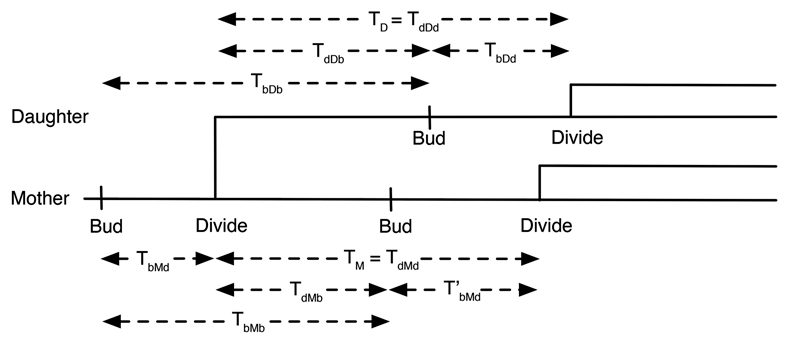
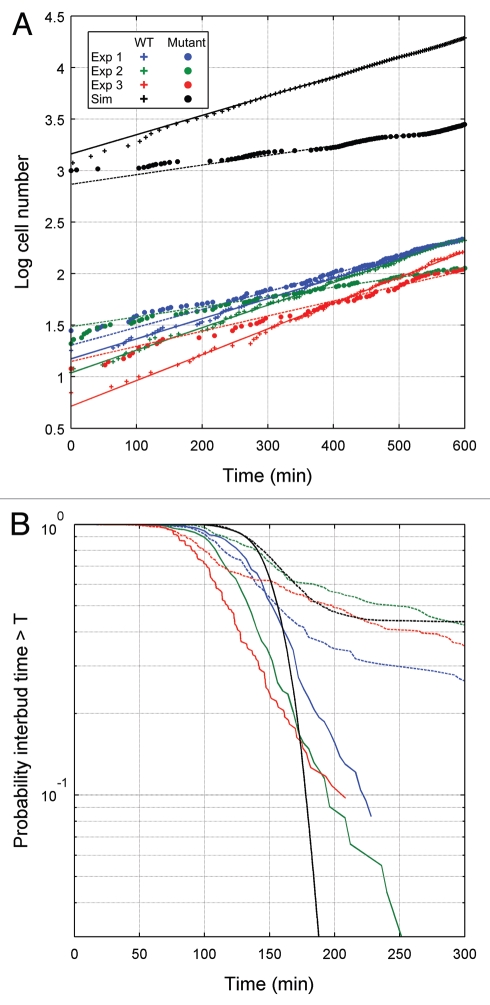
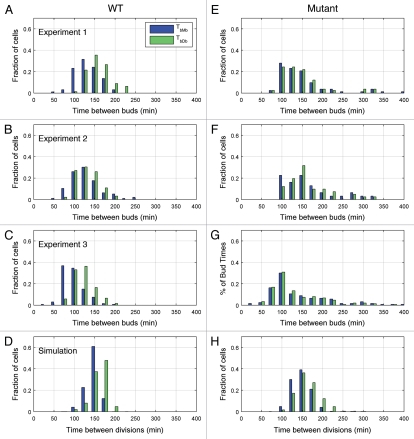
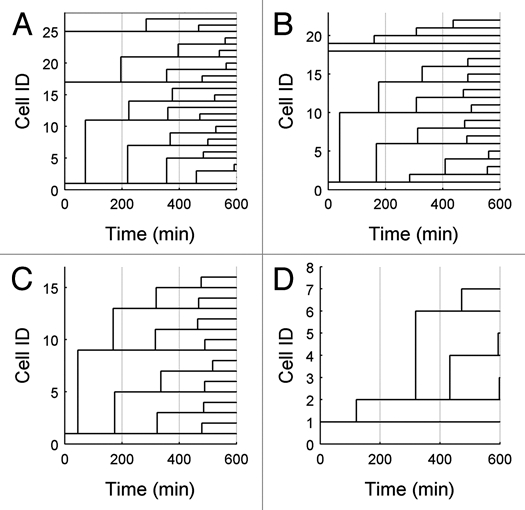
Unlike many mutants that are completely viable or inviable, the CLB2-dbΔ clb5Δ mutant of Saccharomyces cerevisiae is inviable in glucose but partially viable on slower growth media such as raffinose. On raffinose, the mutant cells can bud and divide but in each cycle there is a chance that a cell will fail to divide (telophase arrest), causing it to exit the cell cycle. This effect gives rise to a stochastic phenotype that cannot be explained by a deterministic model. We measure the inter-bud times of wild type and mutant cells growing on raffinose and compute statistics and distributions to characterize the mutant's behavior. We convert a detailed deterministic model of the budding yeast cell cycle to a stochastic model and determine the extent to which it captures the stochastic phenotype of the mutant strain. Predictions of the mathematical model are in reasonable agreement with our experimental data and suggest directions for improving the model. Ultimately, the ability to accurately model stochastic phenotypes may prove critical to understanding disease and therapeutic interventions in higher eukaryotes.
Figures
Comment in
-
Two legs are better than one.Cell Cycle. 2011 Apr 15;10(8):1189-90. doi: 10.4161/cc.10.8.15298. Epub 2011 Apr 15. Cell Cycle. 2011. PMID: 21436620 No abstract available.
References
-
- Weinberg RA. The biology of cancer. London: Garland Science; 2006.
-
- Lorincz A, Carter BLA. Control of cell size at bud initiation in Saccharomyces cerevisiae. Journal of General Microbiology. 1979;113:287–295.
-
- Sinclair DA, Mills K, Guarente L. Molecular mechanisms of yeast aging. Trends Biochem Sci. 1998;23:131–134. - PubMed
-
- Wäsch R, Cross FR. Apc-dependent proteolysis of the mitotic cyclin clb2 is essential for mitotic exit. Nature. 2002;418:556–562. - PubMed
-
- Cross FR. Two redundant oscillatory mechanisms in the yeast cell cycle. Dev Cell. 2003;4:741–752. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases