

Direct binding of Cenp-C to the Mis12 complex joins the inner and outer kinetochore
- PMID: 21353556
- PMCID: PMC3074538
- DOI: 10.1016/j.cub.2010.12.039
Direct binding of Cenp-C to the Mis12 complex joins the inner and outer kinetochore
Abstract
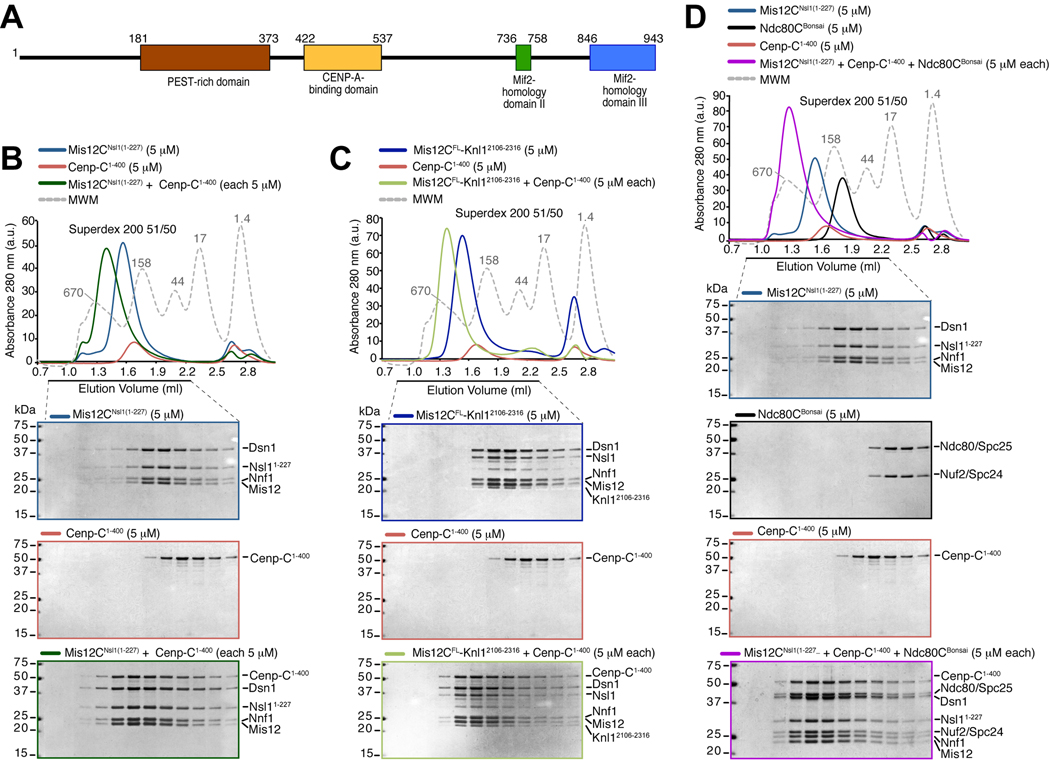
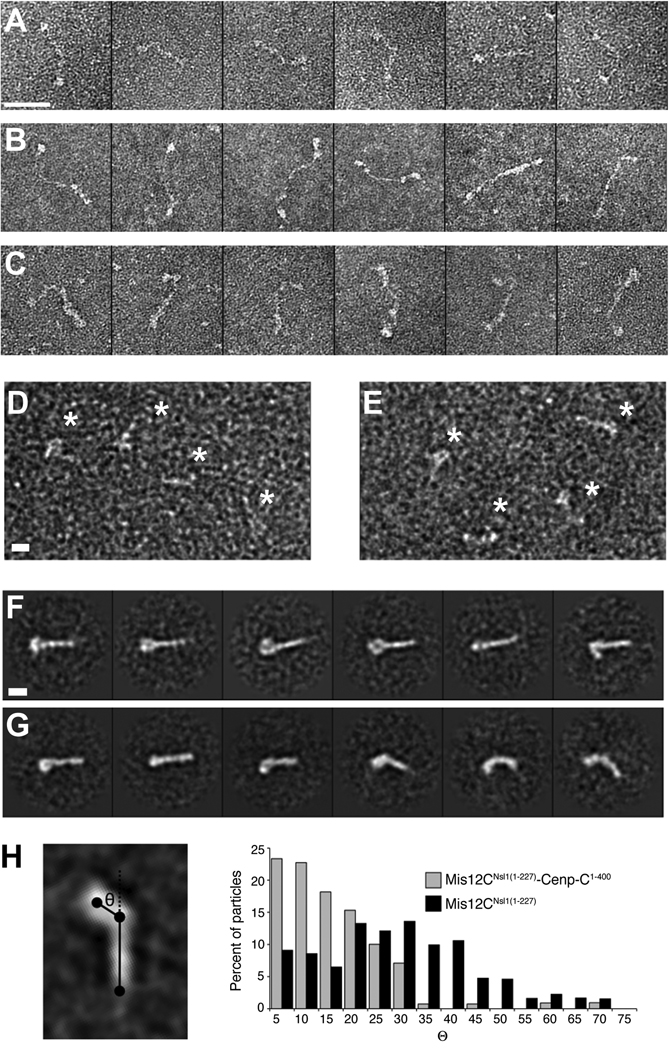
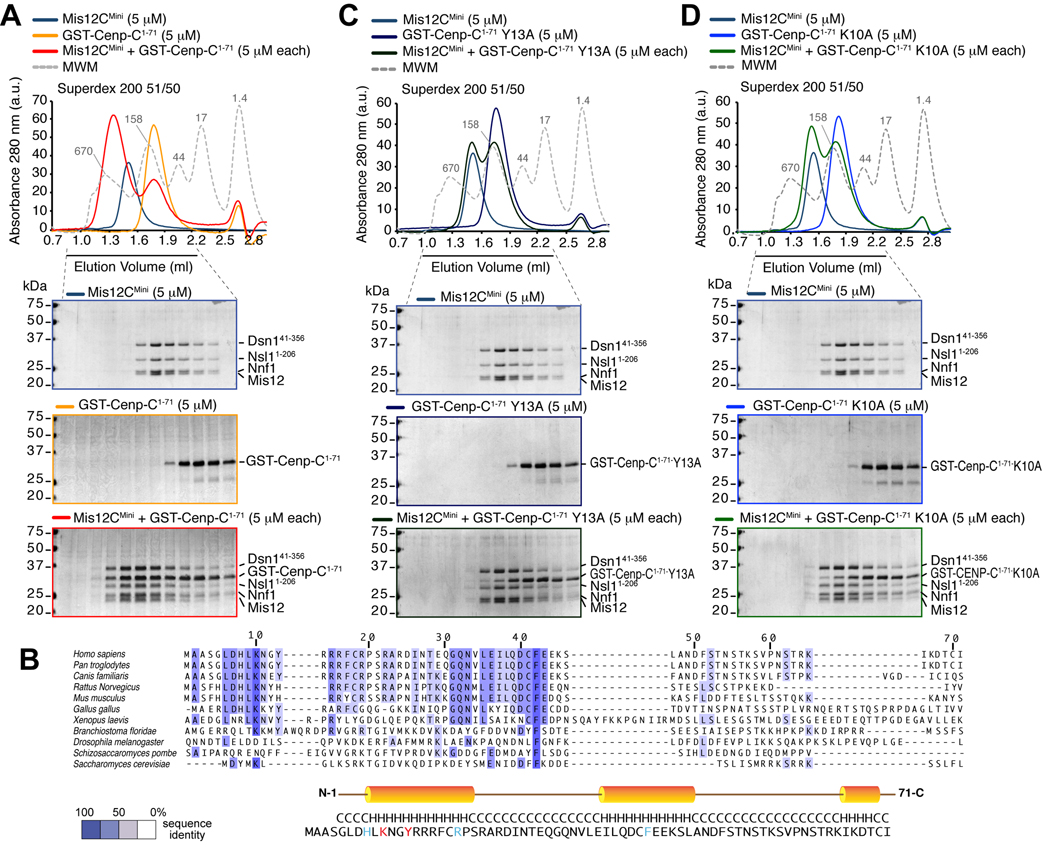
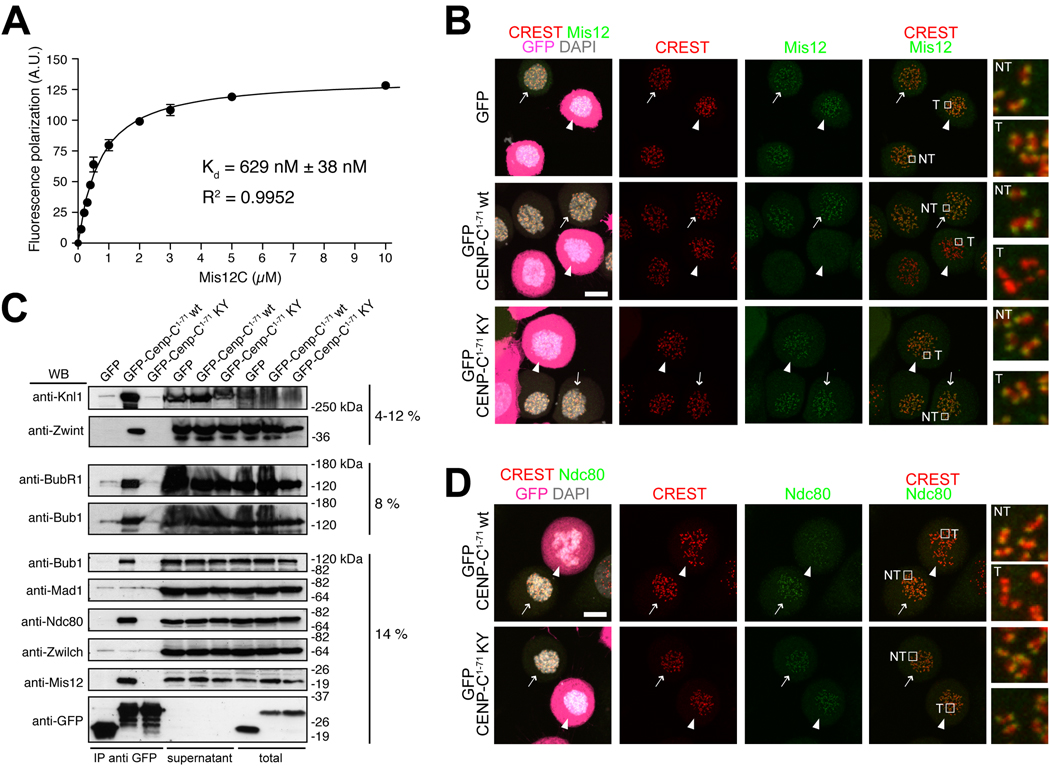
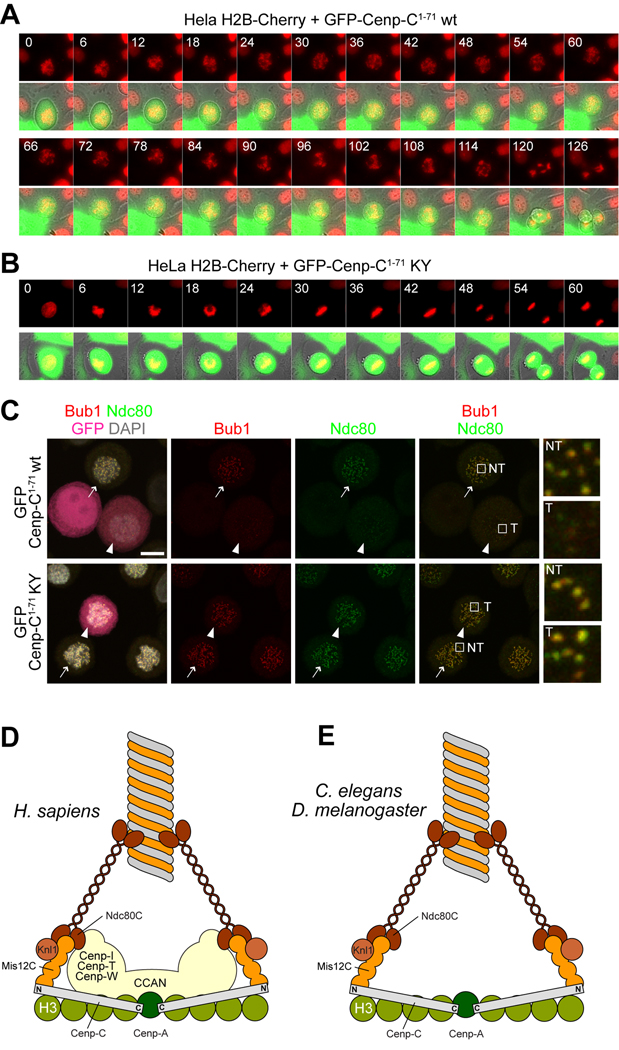
Kinetochores are proteinaceous scaffolds implicated in the formation of load-bearing attachments of chromosomes to microtubules during mitosis. Kinetochores contain distinct chromatin- and microtubule-binding interfaces, generally defined as the inner and outer kinetochore, respectively (reviewed in). The constitutive centromere-associated network (CCAN) and the Knl1-Mis12-Ndc80 complexes (KMN) network are the main multisubunit protein assemblies in the inner and outer kinetochore, respectively. The point of contact between the CCAN and the KMN network is unknown. Cenp-C is a conserved CCAN component whose central and C-terminal regions have been implicated in chromatin binding and dimerization. Here, we show that a conserved motif in the N-terminal region of Cenp-C binds directly and with high affinity to the Mis12 complex. Expression in HeLa cells of the isolated N-terminal motif of Cenp-C prevents outer kinetochore assembly, causing chromosome missegregation. The KMN network is also responsible for kinetochore recruitment of the components of the spindle assembly checkpoint, and we observe checkpoint impairment in cells expressing the Cenp-C N-terminal segment. Our studies unveil a crucial and likely universal link between the inner and outer kinetochore.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Chromosome segregation: a kinetochore missing link is found.Curr Biol. 2011 Apr 12;21(7):R261-3. doi: 10.1016/j.cub.2011.02.026. Curr Biol. 2011. PMID: 21481764
References
-
- Trazzi S, Bernardoni R, Diolaiti D, Politi V, Earnshaw WC, Perini G, Della Valle G. In vivo functional dissection of human inner kinetochore protein CENP-C. J Struct Biol. 2002;140:39–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
