

A sensory code for host seeking in parasitic nematodes
- PMID: 21353558
- PMCID: PMC3152378
- DOI: 10.1016/j.cub.2011.01.048
A sensory code for host seeking in parasitic nematodes
Abstract
Parasitic nematode species often display highly specialized host-seeking behaviors that reflect their specific host preferences. Many such behaviors are triggered by host odors, but little is known about either the specific olfactory cues that trigger these behaviors or the underlying neural circuits. Heterorhabditis bacteriophora and Steinernema carpocapsae are phylogenetically distant insect-parasitic nematodes whose host-seeking and host-invasion behavior resembles that of some devastating human- and plant-parasitic nematodes. We compare the olfactory responses of Heterorhabditis and Steinernema infective juveniles (IJs) to those of Caenorhabditis elegans dauers, which are analogous life stages. The broad host range of these parasites results from their ability to respond to the universally produced signal carbon dioxide (CO(2)), as well as a wide array of odors, including host-specific odors that we identified using thermal desorption-gas chromatography-mass spectroscopy. We find that CO(2) is attractive for the parasitic IJs and C. elegans dauers despite being repulsive for C. elegans adults, and we identify a sensory neuron that mediates CO(2) response in both parasitic and free-living species, regardless of whether CO(2) is attractive or repulsive. The parasites' odor response profiles are more similar to each other than to that of C. elegans despite their greater phylogenetic distance, likely reflecting evolutionary convergence to insect parasitism.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
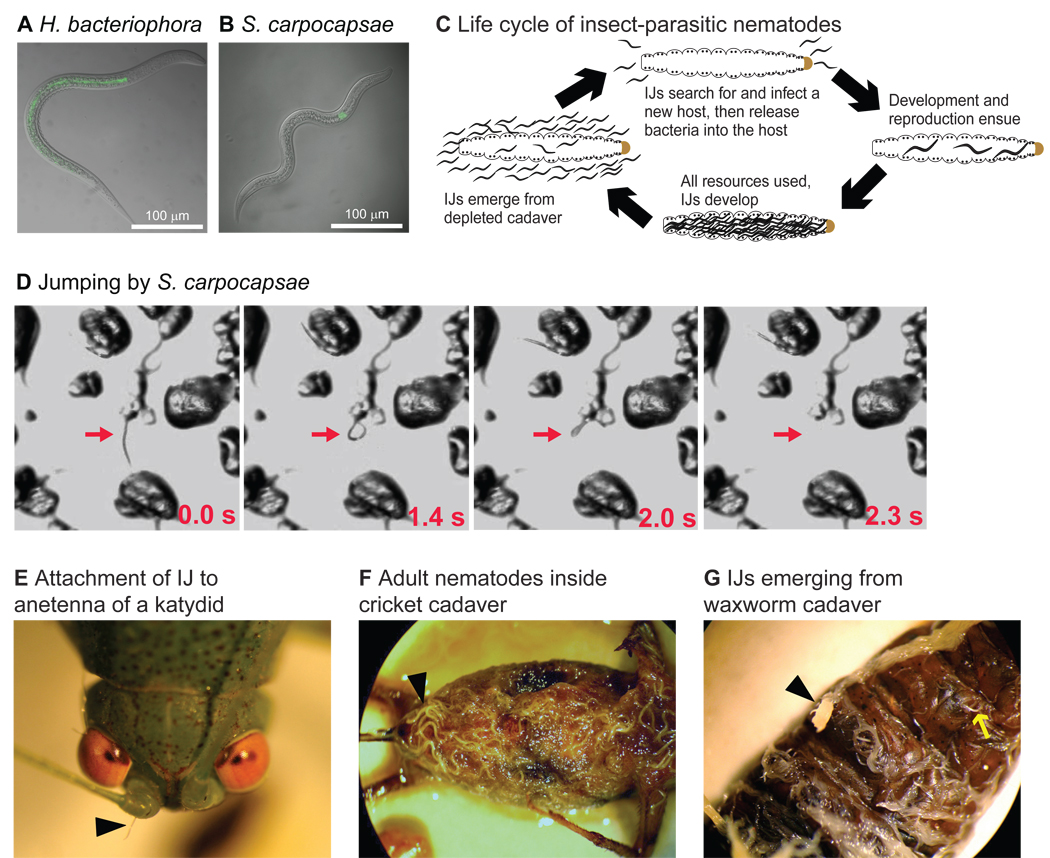
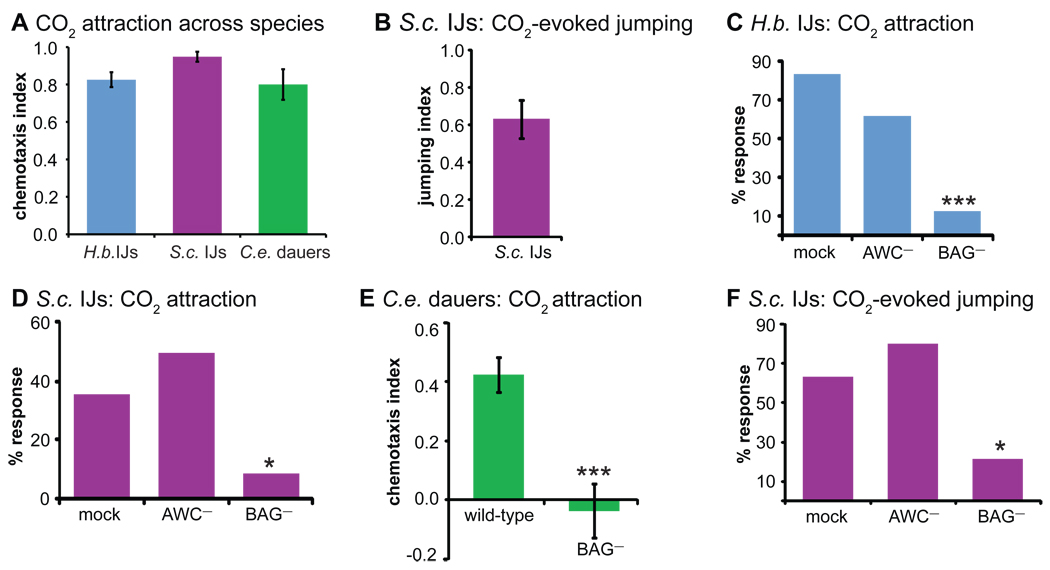
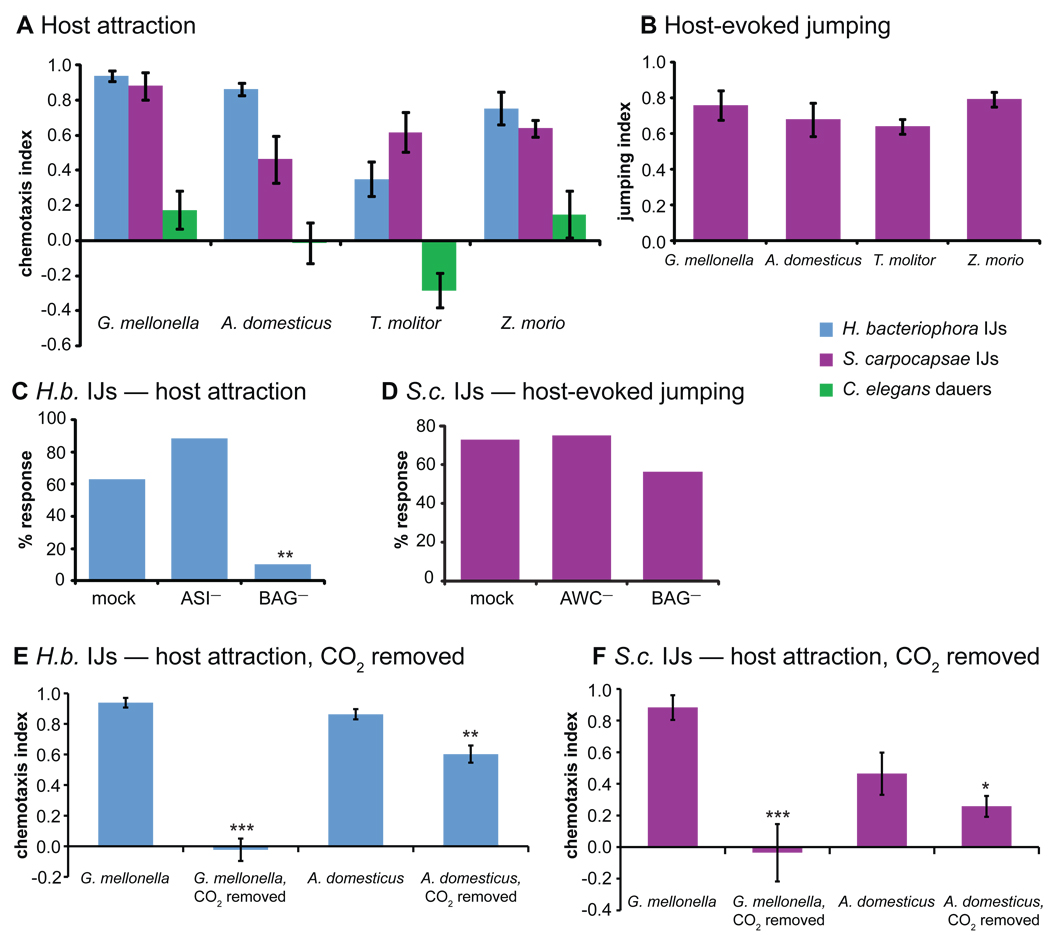
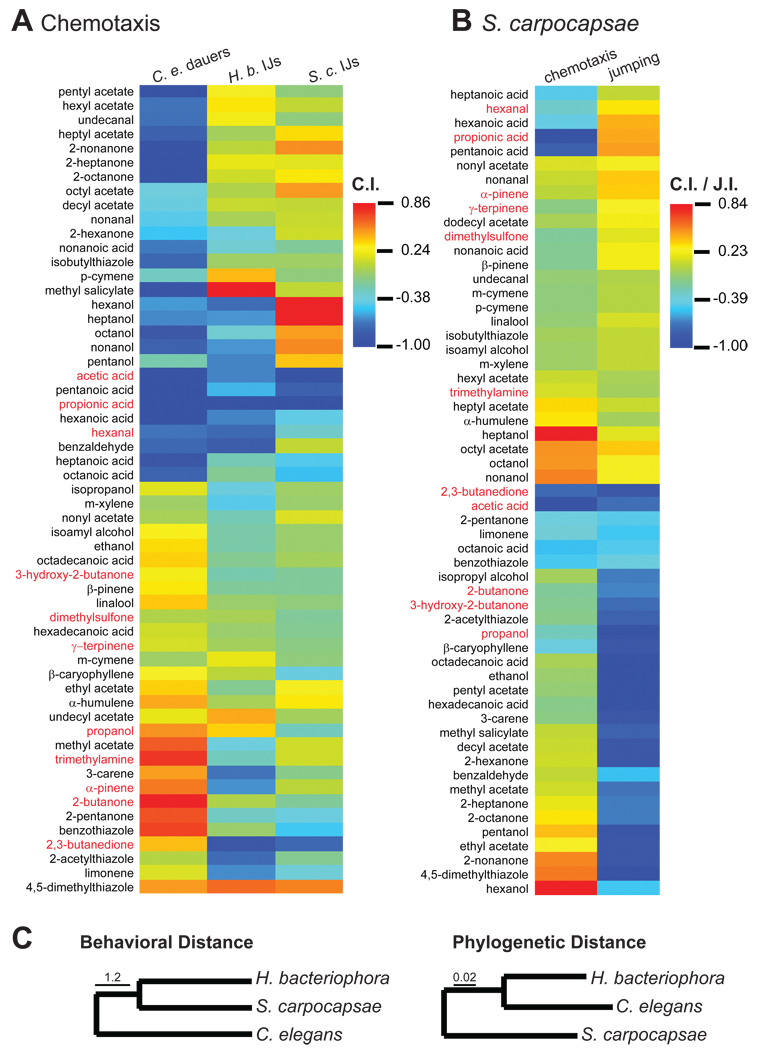
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
