

MSUT2 is a determinant of susceptibility to tau neurotoxicity
- PMID: 21355046
- PMCID: PMC3080609
- DOI: 10.1093/hmg/ddr079
MSUT2 is a determinant of susceptibility to tau neurotoxicity
Abstract
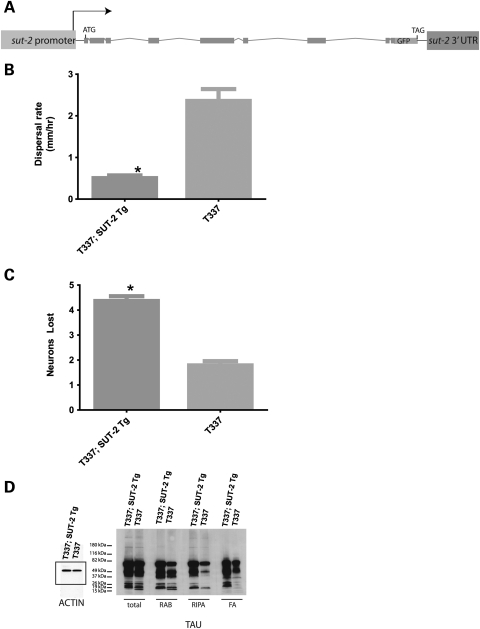
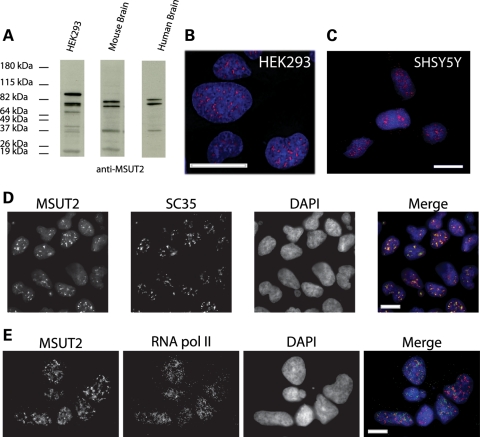
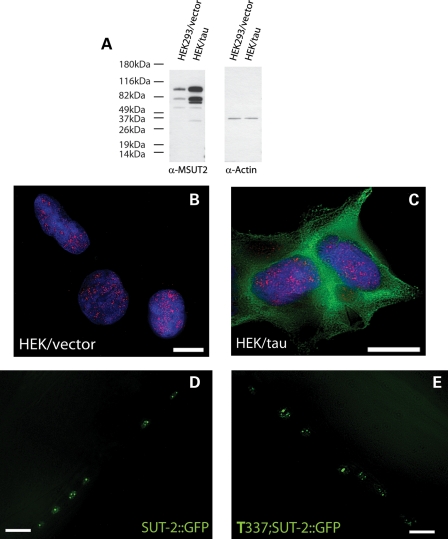
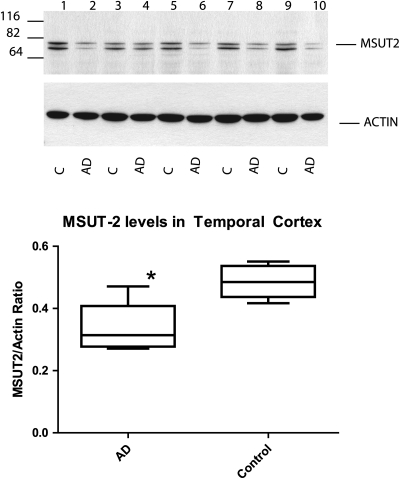
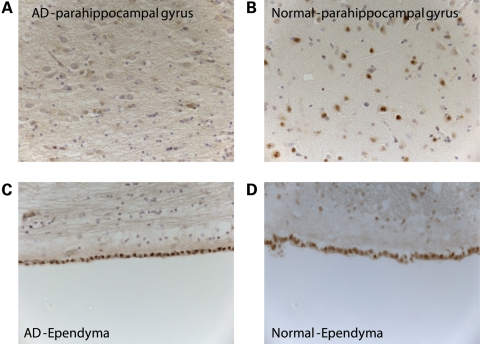
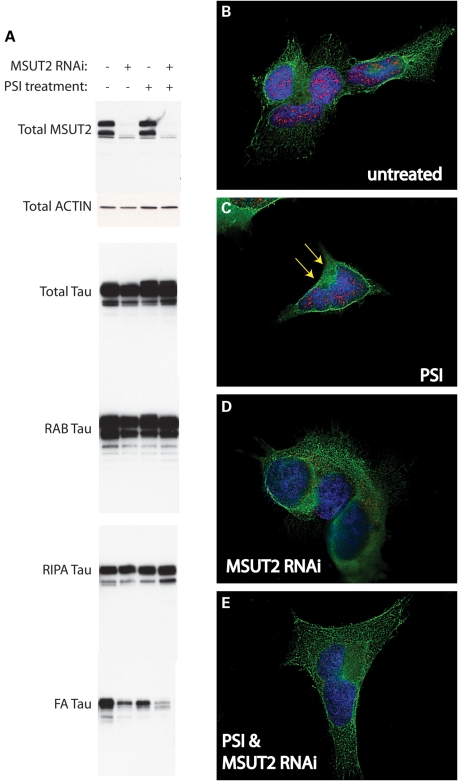
Lesions containing abnormal aggregated tau protein are one of the diagnostic hallmarks of Alzheimer's disease (AD) and related tauopathy disorders. How aggregated tau leads to dementia remains enigmatic, although neuronal dysfunction and loss clearly contribute. We previously identified sut-2 as a gene required for tau neurotoxicity in a transgenic Caenorhabditis elegans model of tauopathy. Here, we further explore the role of sut-2 and show that overexpression of SUT-2 protein enhances tau-induced neuronal dysfunction, neurotoxicity and accumulation of insoluble tau. We also explore the relationship between sut-2 and its human homolog, mammalian SUT-2 (MSUT2) and find both proteins to be predominantly nuclear and localized to SC35-positive nuclear speckles. Using a cell culture model for the accumulation of pathological tau, we find that high tau levels lead to increased expression of MSUT2 protein. We analyzed MSUT2 protein in age-matched post-mortem brain samples from AD patients and observe a marked decrease in overall MSUT2 levels in the temporal lobe of AD patients. Analysis of post-mortem tissue from AD cases shows a clear reduction in neuronal MSUT2 levels in brain regions affected by tau pathology, but little change in regions lacking tau pathology. RNAi knockdown of MSUT2 in cultured human cells overexpressing tau causes a marked decrease in tau aggregation. Both cell culture and post-mortem tissue studies suggest that MSUT2 levels may influence neuronal vulnerability to tau toxicity and aggregation. Thus, neuroprotective strategies targeting MSUT2 may be of therapeutic interest for tauopathy disorders.
Figures
References
-
- Schneider A., Mandelkow E. Tau-based treatment strategies in neurodegenerative diseases. Neurotherapeutics. 2008;5:443–457. doi:10.1016/j.nurt.2008.05.006. - DOI - PMC - PubMed
-
- Kertesz A. Clinical features and diagnosis of frontotemporal dementia. Front. Neurol. Neurosci. 2009;24:140–148. doi:10.1159/000197893. - DOI - PubMed
-
- McKee A.C., Cantu R.C., Nowinski C.J., Hedley-Whyte E.T., Gavett B.E., Budson A.E., Santini V.E., Lee H.S., Kubilus C.A., Stern R.A. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J. Neuropathol. Exp. Neurol. 2009;68:709–735. doi:10.1097/NEN.0b013e3181a9d503. - DOI - PMC - PubMed
-
- Spillantini M.G., Murrell J.R., Goedert M., Farlow M.R., Klug A., Ghetti B. Mutation in the tau gene in familial multiple system tauopathy with presenile dementia. Proc. Natl Acad. Sci. USA. 1998;95:7737–7741. doi:10.1073/pnas.95.13.7737. - DOI - PMC - PubMed
-
- Clark L.N., Poorkaj P., Wszolek Z., Geschwind D.H., Nasreddine Z.S., Miller B., Li D., Payami H., Awert F., Markopoulou K., et al. Pathogenic implications of mutations in the tau gene in pallido-ponto-nigral degeneration and related neurodegenerative disorders linked to chromosome 17. Proc. Natl Acad. Sci. USA. 1998;95:13103–13107. doi:10.1073/pnas.95.22.13103. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
