

Review
doi: 10.1016/j.cmet.2011.02.004.
Regulation of lipid homeostasis by the bifunctional SREBF2-miR33a locus
Affiliations
- PMID: 21356514
- PMCID: PMC3062104
- DOI: 10.1016/j.cmet.2011.02.004
Item in Clipboard
Review
Regulation of lipid homeostasis by the bifunctional SREBF2-miR33a locus
Cell Metab.
.
Abstract
The sterol regulatory element-binding factor-2 (SREBF2) gene is a bifunctional locus encoding SREBP-2, a well-known transcriptional regulator of genes involved in cholesterol biosynthesis, and microRNA-33a, which has recently been shown to reduce expression of proteins involved in export of cholesterol and β-oxidation of fatty acids, thus adding an unexpected layer of complexity and fine-tuning to regulation of lipid homeostasis.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
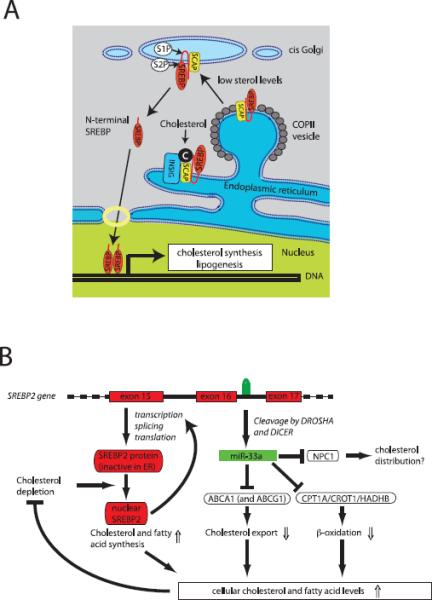
(A) Traditional model of SREBP-2 function. SREBP-2 contains two transmembrane domains and is inserted in the membrane of the endoplasmic reticulum (ER). Under conditions of abundant cholesterol (or phosphatidylethanolamine), SREBP2 is retained in the ER by a complex of SCAP and INSIG proteins. When cholesterol levels fall, the interaction between INSIG and SCAP is disrupted, allowing SCAP to interact with COPII-coated vesicles and target SREBP-2 for export to the Golgi apparatus. Here, in two consecutive cleavages by S1P and S2P, a transcriptionally competent N-terminal fragment is released. After translocation to the nucleus, SREBP-2 induces expression of genes involved in cholesterol and fatty acid synthesis, which serve to normalize cellular lipid levels (modified from (Ikonen, 2008)). (B) Integrated model for how the bifunctional SREBF2 locus maintains lipid homeostasis. After processing from an intron of SREBF2, miR-33a reduces cellular cholesterol export by inhibiting expression of ABCA1 (and in the mouse ABCG1). In addition, miR-33a reduces mitochondrial fatty acid β-oxidation via inhibition of HADHB, CROT and CPT1A to increase intracellular lipid levels. Thus the SREBF2 locus uses two distinct mechanisms to maintain lipid homeostasis: regulated transcriptional activity of SREBP-2 and translational repression by miR-33a.

miR-33a is generated from an intron of the SREBF2 primary transcript and inhibits translation of ABCA1 and ABCG1 mRNAs, and perhaps NPC1. Putative flux of cholesterol is denoted with red arrows under conditions of A) low sterol concentrations (A) or high sterol concentrations/miR-33a deficiency (B). De novo HDL synthesis requires ABCA1-dependent cholesterol secretion from the liver and intestine into nascent HDL particles. In peripheral organs, HDL-particles accept cholesterol from ABCA1 and ABCG1 transporters. Hepatic uptake of HDL particles via the scavenger receptor-B1 (SR-B1) completes the process of reverse cholesterol transport. To deliver fatty acids and cholesterol to peripheral organs, the liver synthesizes VLDL particles. Delivery of fatty acids and cholesterol eventually leads to the transformation of VLDL into LDL particles, which are cleared from circulation via hepatic LDL receptor (LDLR). Oxidation and/or impaired clearance of LDL increase risk of foam cell formation and atheroma formation. Upon reduction of hepatic cholesterol levels, SREBP-2 increases transcription of genes involved in cholesterol synthesis (e.g. β-HMG CoA reductase), as well as LDLR to increase uptake of LDL. (A) Low sterol concentraions promote the expression of miR-33a, which inhibits the expression of ABCA1 and ABCG1 to suppress cholesterol export from liver, intestine and macrophages and thereby prevents further decreases in intercellular sterol concentrations. miR-33a may also influence intercellular cholesterol distribution by inhibiting NPC1 expression. (B) Suppression of miR-33a expression by either high sterol concentrations or experimental intervention results in increased ABCA1 and ABCG1 expression, which potentially increases not only de novo synthesis of HDL particles but also loading with cholesterol in the periphery. For simplicicty, we did not include a schematic representation of SREBP-2 expression in peripheral tissues, and do not show effects of cholesterol synthesis and cholesterol secretion.
Similar articles
-
MicroRNAs regulating lipid metabolism in atherogenesis.Thromb Haemost. 2012 Apr;107(4):642-7. doi: 10.1160/TH11-10-0694. Epub 2012 Jan 25. Thromb Haemost. 2012. PMID: 22274626 Free PMC article. Review.
-
Methyl protodioscin increases ABCA1 expression and cholesterol efflux while inhibiting gene expressions for synthesis of cholesterol and triglycerides by suppressing SREBP transcription and microRNA 33a/b levels.Atherosclerosis. 2015 Apr;239(2):566-70. doi: 10.1016/j.atherosclerosis.2015.02.034. Epub 2015 Feb 23. Atherosclerosis. 2015. PMID: 25733328
-
SREBP-2-deficient and hypomorphic mice reveal roles for SREBP-2 in embryonic development and SREBP-1c expression.J Lipid Res. 2016 Mar;57(3):410-21. doi: 10.1194/jlr.M064022. Epub 2015 Dec 18. J Lipid Res. 2016. PMID: 26685326 Free PMC article.
-
Enhanced Liver Regeneration After Partial Hepatectomy in Sterol Regulatory Element-Binding Protein (SREBP)-1c-Null Mice is Associated with Increased Hepatocellular Cholesterol Availability.Cell Physiol Biochem. 2018;47(2):784-799. doi: 10.1159/000490030. Epub 2018 May 22. Cell Physiol Biochem. 2018. PMID: 29807364
-
Endoplasmic reticulum stress and lipid dysregulation.Expert Rev Mol Med. 2011 Feb 3;13:e4. doi: 10.1017/S1462399410001742. Expert Rev Mol Med. 2011. PMID: 21288373 Review.
Cited by
-
MicroRNA-33a mediates the regulation of high mobility group AT-hook 2 gene (HMGA2) by thyroid transcription factor 1 (TTF-1/NKX2-1).J Biol Chem. 2013 Jun 7;288(23):16348-16360. doi: 10.1074/jbc.M113.474643. Epub 2013 Apr 26. J Biol Chem. 2013. PMID: 23625920 Free PMC article.
-
Consequences of exchanging carbohydrates for proteins in the cholesterol metabolism of mice fed a high-fat diet.PLoS One. 2012;7(11):e49058. doi: 10.1371/journal.pone.0049058. Epub 2012 Nov 6. PLoS One. 2012. PMID: 23139832 Free PMC article.
-
Beneficial effects of cherry consumption as a dietary intervention for metabolic, hepatic and vascular complications in type 2 diabetic rats.Cardiovasc Diabetol. 2018 Jul 20;17(1):104. doi: 10.1186/s12933-018-0744-6. Cardiovasc Diabetol. 2018. PMID: 30029691 Free PMC article.
-
Interaction effect between NAFLD severity and high carbohydrate diet on gut microbiome alteration and hepatic de novo lipogenesis.Gut Microbes. 2022 Jan-Dec;14(1):2078612. doi: 10.1080/19490976.2022.2078612. Gut Microbes. 2022. PMID: 35634707 Free PMC article.
-
A global view of aging and Alzheimer's pathogenesis-associated cell population dynamics and molecular signatures in human and mouse brains.Nat Genet. 2023 Dec;55(12):2104-2116. doi: 10.1038/s41588-023-01572-y. Epub 2023 Nov 30. Nat Genet. 2023. PMID: 38036784 Free PMC article.
References
-
- Brunham LR, Kastelein JJ, Hayden MR. ABCA1 gene mutations, HDL cholesterol levels, and risk of ischemic heart disease. JAMA. 2008;300:1997–1998. author reply 1998. - PubMed
-
- Brunham LR, Singaraja RR, Duong M, Timmins JM, Fievet C, Bissada N, Kang MH, Samra A, Fruchart JC, McManus B, Staels B, Parks JS, Hayden MR. Tissue-specific roles of ABCA1 influence susceptibility to atherosclerosis. Arterioscler Thromb Vasc Biol. 2009;29:548–554. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
