

Transcriptional control of adipose lipid handling by IRF4
- PMID: 21356515
- PMCID: PMC3063358
- DOI: 10.1016/j.cmet.2011.02.005
Transcriptional control of adipose lipid handling by IRF4
Abstract
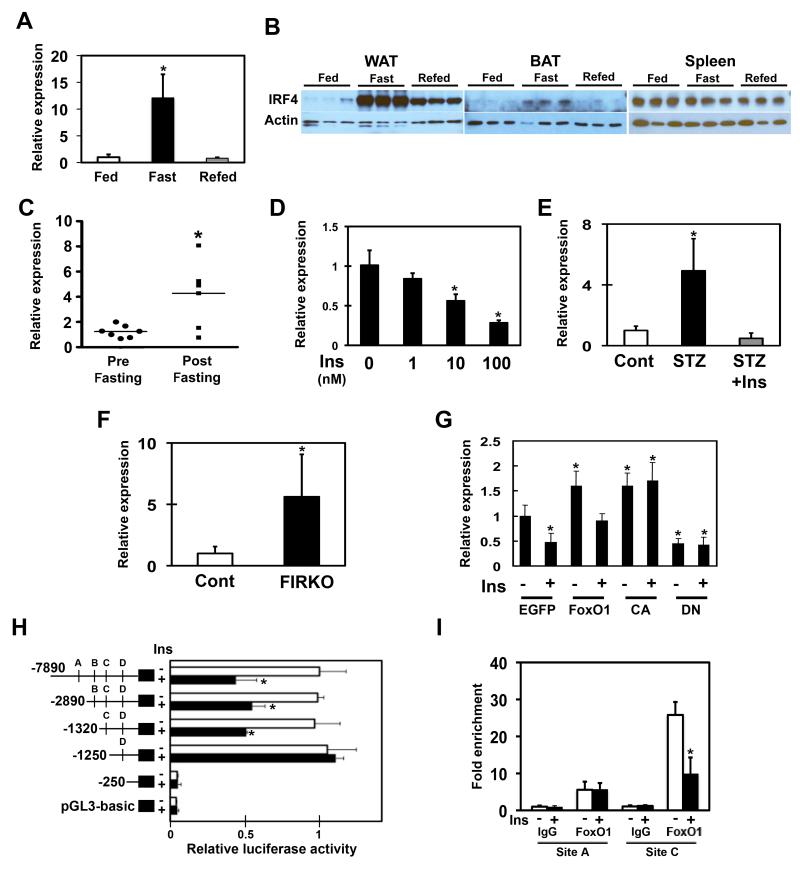
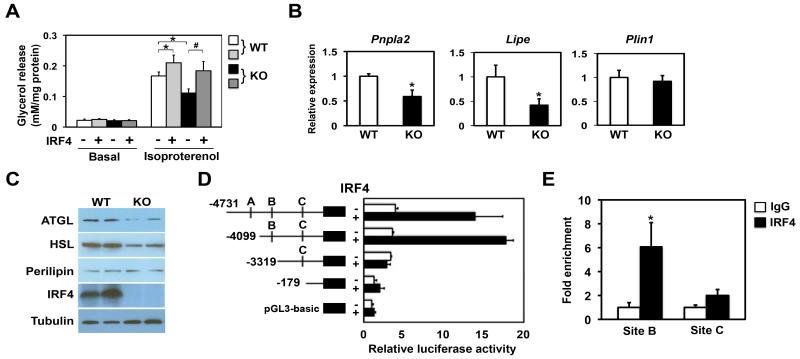
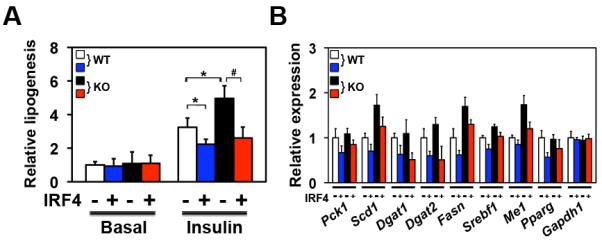
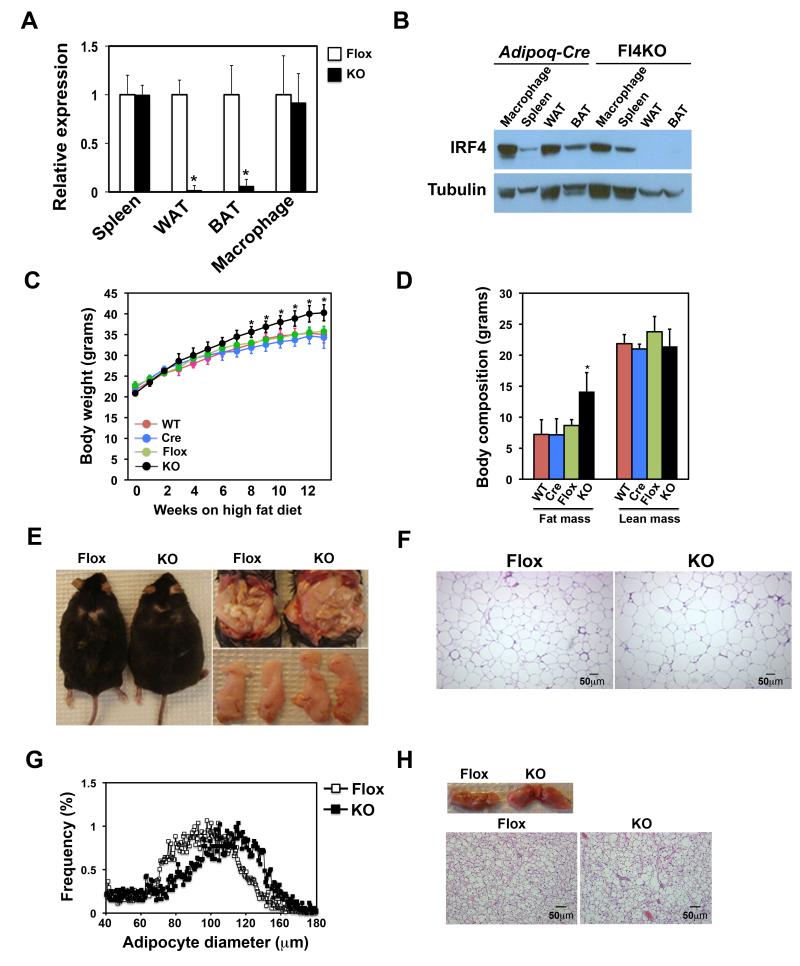
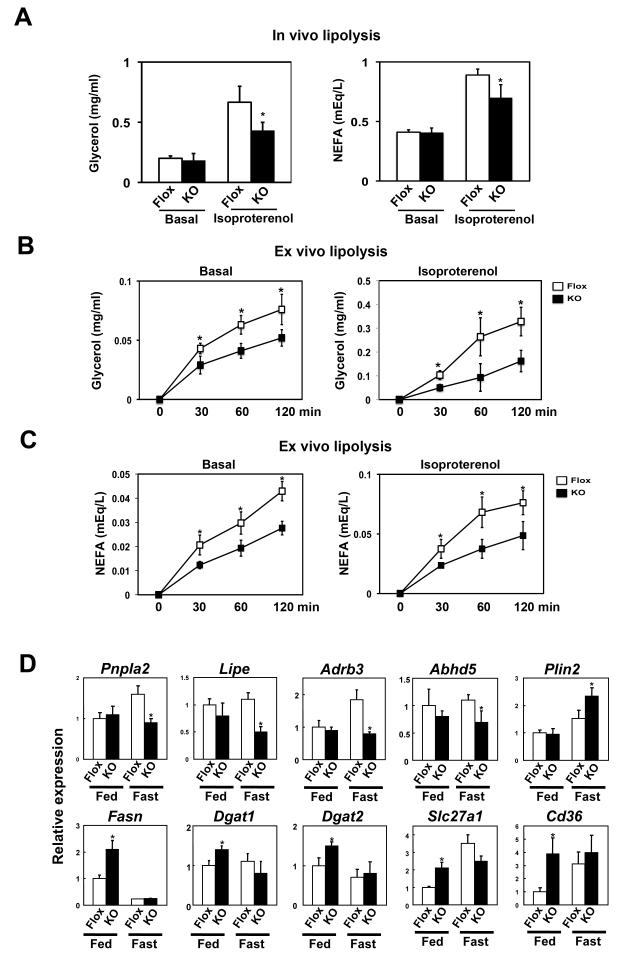
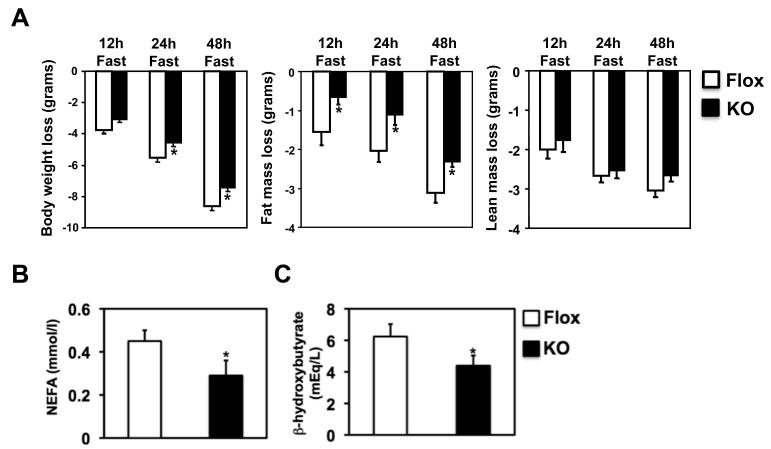
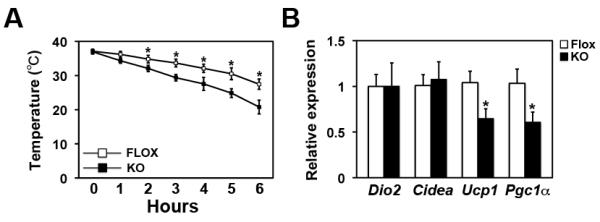
Adipocytes store triglyceride during periods of nutritional affluence and release free fatty acids during fasting through coordinated cycles of lipogenesis and lipolysis. While much is known about the acute regulation of these processes during fasting and feeding, less is understood about the transcriptional basis by which adipocytes control lipid handling. Here, we show that interferon regulatory factor 4 (IRF4) is a critical determinant of the transcriptional response to nutrient availability in adipocytes. Fasting induces IRF4 in an insulin- and FoxO1-dependent manner. IRF4 is required for lipolysis, at least in part due to direct effects on the expression of adipocyte triglyceride lipase and hormone-sensitive lipase. Conversely, reduction of IRF4 enhances lipid synthesis. Mice lacking adipocyte IRF4 exhibit increased adiposity and deficient lipolysis. These studies establish a link between IRF4 and the disposition of calories in adipose tissue, with consequences for systemic metabolic homeostasis.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Abel ED, Peroni O, Kim JK, Kim YB, Boss O, Hadro E, Minnemann T, Shulman GI, Kahn BB. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature. 2001;409:729–733. - PubMed
-
- Barlow C, Schroeder M, Lekstrom-Himes J, Kylefjord H, Deng CX, Wynshaw-Boris A, Spiegelman BM, Xanthopoulos KG. Targeted expression of Cre recombinase to adipose tissue of transgenic mice directs adipose-specific excision of loxP-flanked gene segments. Nucleic Acids Res. 1997;25:2543–2545. - PMC - PubMed
-
- Bluher M. Adipose tissue dysfunction in obesity. Exp Clin Endocrinol Diabetes. 2009;117:241–250. - PubMed
-
- Bluher M, Michael MD, Peroni OD, Ueki K, Carter N, Kahn BB, Kahn CR. Adipose tissue selective insulin receptor knockout protects against obesity and obesity-related glucose intolerance. Dev Cell. 2002;3:25–38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
