

Electrophilic nitro-fatty acids activate NRF2 by a KEAP1 cysteine 151-independent mechanism
- PMID: 21357422
- PMCID: PMC3077602
- DOI: 10.1074/jbc.M110.190710
Electrophilic nitro-fatty acids activate NRF2 by a KEAP1 cysteine 151-independent mechanism
Abstract
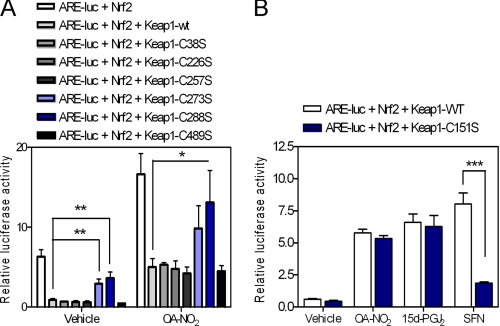
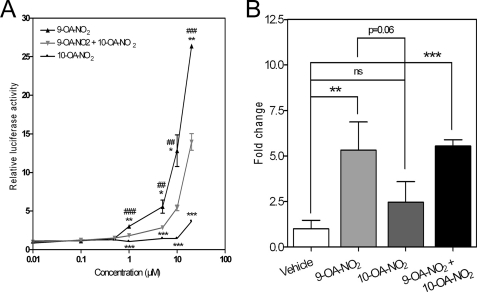
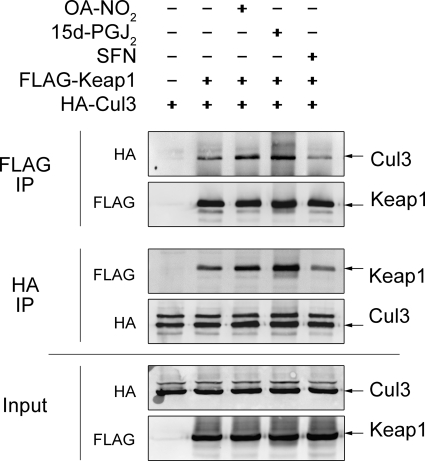
Nitro-fatty acids (NO(2)-FAs) are electrophilic signaling mediators formed in vivo via nitric oxide (NO)- and nitrite (NO(2)(-))-dependent reactions. Nitro-fatty acids modulate signaling cascades via reversible covalent post-translational modification of nucleophilic amino acids in regulatory proteins and enzymes, thus altering downstream signaling events, such as Keap1-Nrf2-antioxidant response element (ARE)-regulated gene expression. In this study, we investigate the molecular mechanisms by which 9- and 10-nitro-octadec-9-enoic acid (OA-NO(2)) activate the transcription factor Nrf2, focusing on the post-translational modifications of cysteines in the Nrf2 inhibitor Keap1 by nitroalkylation and its downstream responses. Of the two regioisomers, 9-nitro-octadec-9-enoic acid was a more potent ARE inducer than 10-nitro-octadec-9-enoic acid. The most OA-NO(2)-reactive Cys residues in Keap1 were Cys(38), Cys(226), Cys(257), Cys(273), Cys(288), and Cys(489). Of these, Cys(273) and Cys(288) accounted for ∼50% of OA-NO(2) reactions in a cellular milieu. Notably, Cys(151) was among the least OA-NO(2)-reactive of the Keap1 Cys residues, with mutation of Cys(151) having no effect on net OA-NO(2) reaction with Keap1 or on ARE activation. Unlike many other Nrf2-activating electrophiles, OA-NO(2) enhanced rather than diminished the binding between Keap1 and the Cul3 subunit of the E3 ligase for Nrf2. OA-NO(2) can therefore be categorized as a Cys(151)-independent Nrf2 activator, which in turn can influence the pattern of gene expression and therapeutic actions of nitroalkenes.
Figures

References
-
- Schopfer F. J., Cole M. P., Groeger A. L., Chen C. S., Khoo N. K., Woodcock S. R., Golin-Bisello F., Motanya U. N., Li Y., Zhang J., Garcia-Barrio M. T., Rudolph T. K., Rudolph V., Bonacci G., Baker P. R., Xu H. E., Batthyany C. I., Chen Y. E., Hallis T. M., Freeman B. A. (2010) J. Biol. Chem. 285, 12321–12333 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
