

Transport and metabolism of the endogenous auxin precursor indole-3-butyric acid
- PMID: 21357648
- PMCID: PMC3098716
- DOI: 10.1093/mp/ssr006
Transport and metabolism of the endogenous auxin precursor indole-3-butyric acid
Abstract
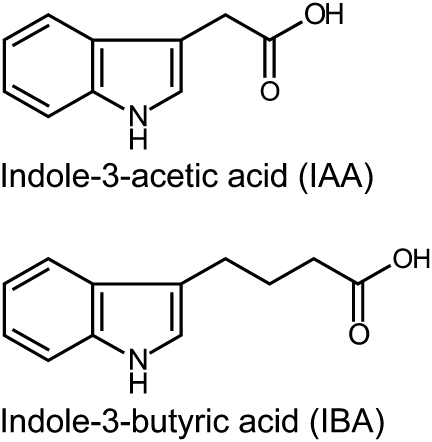
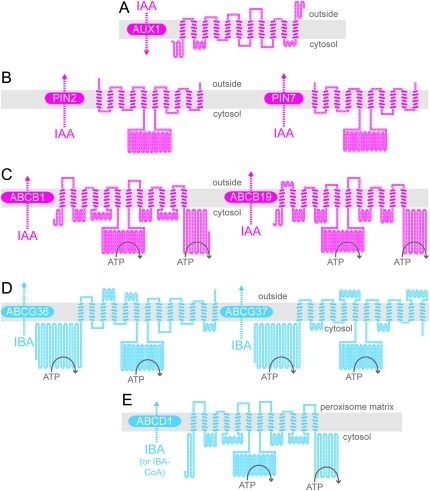
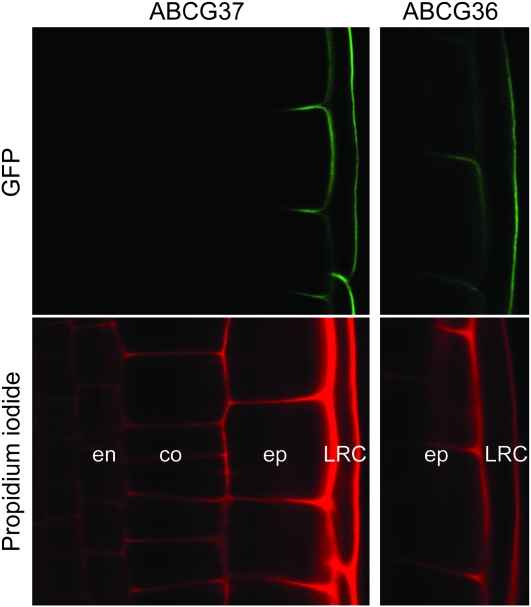
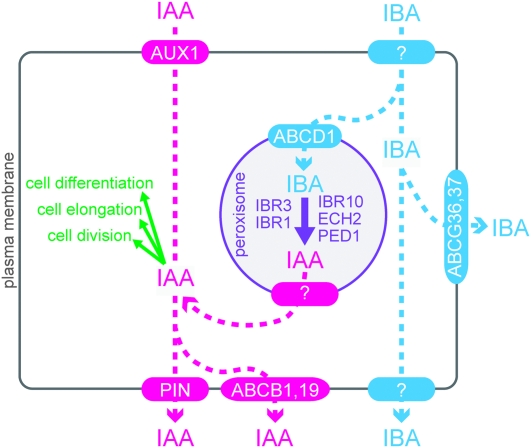
Plant growth and morphogenesis depend on the levels and distribution of the plant hormone auxin. Plants tightly regulate cellular levels of the active auxin indole-3-acetic acid (IAA) through synthesis, inactivation, and transport. Although the transporters that move IAA into and out of cells are well characterized and play important roles in development, little is known about the transport of IAA precursors. In this review, we discuss the accumulating evidence suggesting that the IAA precursor indole-3-butyric acid (IBA) is transported independently of the characterized IAA transport machinery along with the recent identification of specific IBA efflux carriers and enzymes suggested to metabolize IBA. These studies have revealed important roles for IBA in maintaining IAA levels and distribution within the plant to support normal development.
Figures
References
-
- Bailly A, Sovero V, Vincenzetti V, Santelia D, Bartnik D, Koenig BW, Mancuso S, Martinoia E, Geisler M. Modulation of P-glycoproteins by auxin transport inhibitors is mediated by interaction with immunophilins. J. Biol. Chem. 2008;283:21817–21826. - PubMed
-
- Bandyopadhyay A, et al. Interactions of PIN and PGP auxin transport mechanisms. Biochem. Soc. Trans. 2007;35:137–141. - PubMed
-
- Bednarek P, et al. A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science. 2009;323:101–106. - PubMed
-
- Blommaert K. Growth- and inhibiting-substances in relation to the rest period of the potato tuber. Nature. 1954;174:970–972. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
