

Dectin-2 mediates Th2 immunity through the generation of cysteinyl leukotrienes
- PMID: 21357742
- PMCID: PMC3058587
- DOI: 10.1084/jem.20100793
Dectin-2 mediates Th2 immunity through the generation of cysteinyl leukotrienes
Abstract
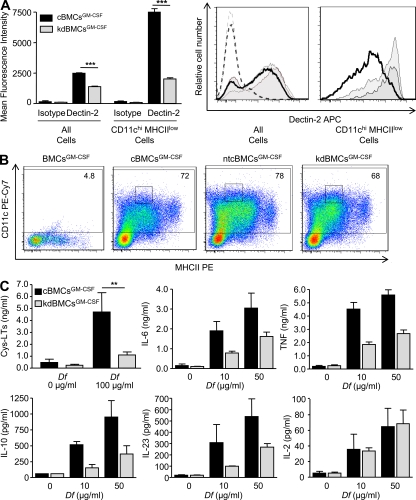
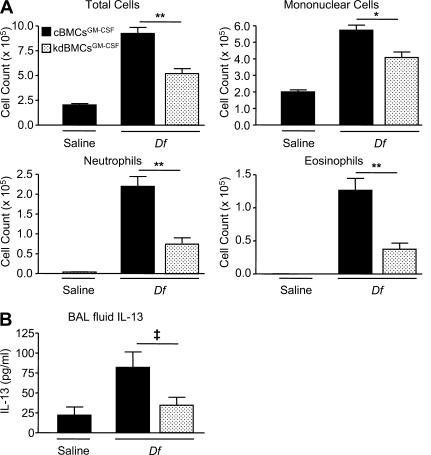
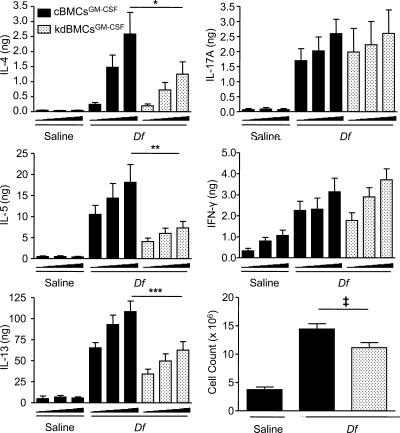
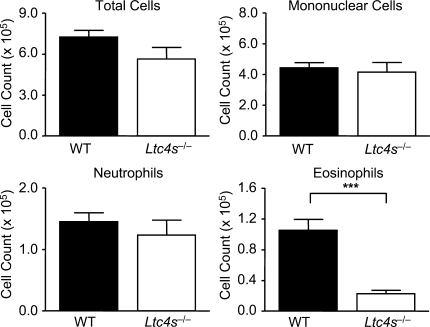
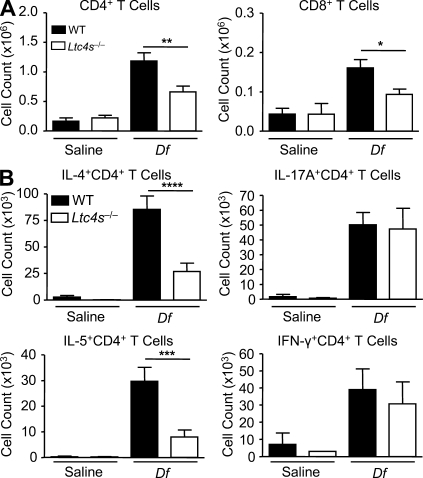
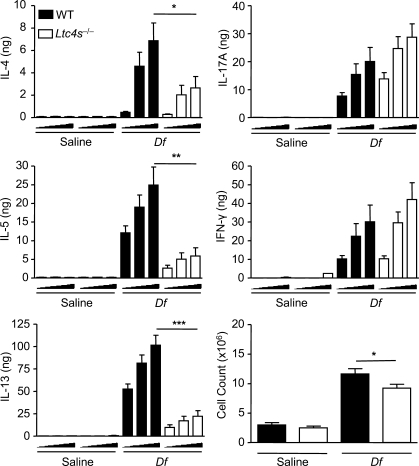
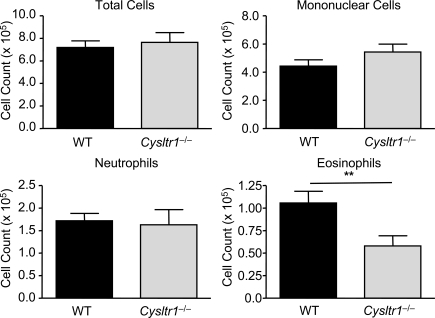
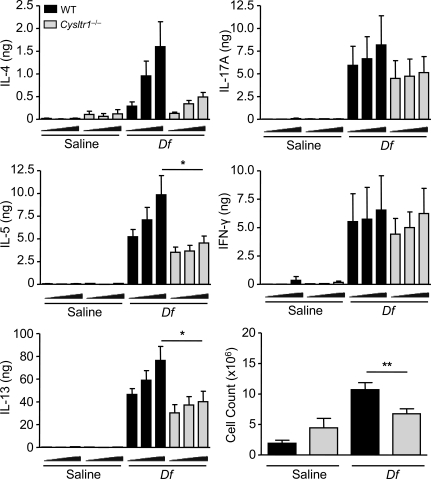
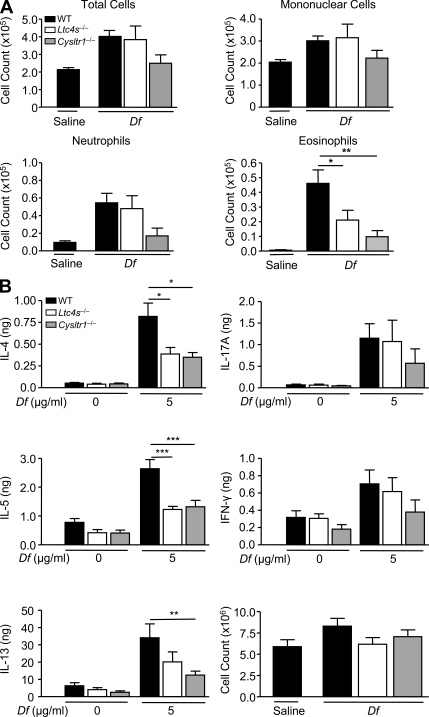
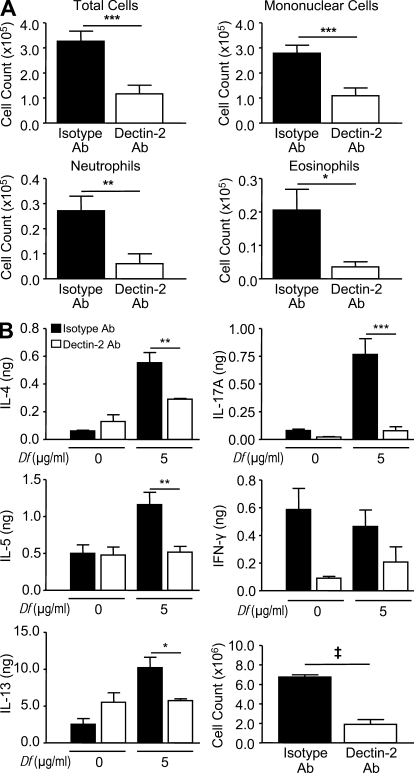
The innate signaling pathways for Th2 immunity activated by inhaled antigens are not well defined. We previously identified Dectin-2 as a receptor for glycans in allergen extracts from the house dust mite Dermatophagoides farinae (Df) that mediates cysteinyl leukotriene (cys-LT) generation from pulmonary CD11c+ cells and from GM-CSF-cultured bone marrow cells (BMCs(GM-CSF)). Using lentiviral knockdown of Dectin-2 in BMCs(GM-CSF) and adoptive transfer of Df-pulsed BMCs(GM-CSF) to sensitize naive mice, we now report that Dectin-2 is critical for the development of Df-elicited eosinophilic and neutrophilic pulmonary inflammation and Th2 cytokine generation in the lungs and restimulated lymph nodes. Sensitization with Df-pulsed BMCs(GM-CSF) from LTC(4) synthase (LTC(4)S)-deficient mice or type 1 cys-LT receptor (CysLT1R)-deficient mice demonstrated that both proteins were required for Df-elicited eosinophilic pulmonary inflammation and Th2 cytokine generation in the lungs and restimulated lymph nodes. Direct sensitization and challenge of Ltc4s-/- and Cysltr1-/- mice confirmed that cys-LTs mediate these parameters of Df-elicited Th2 pulmonary inflammation. Thus, the Dectin-2-cys-LT pathway is critical for the induction of Th2 immunity to a major allergen, in part through CysLT1R. These findings identify a previously unrecognized link between a myeloid C-type lectin receptor and Th2 immunity.
Figures
References
-
- Aragane Y., Maeda A., Schwarz A., Tezuka T., Ariizumi K., Schwarz T. 2003. Involvement of dectin-2 in ultraviolet radiation-induced tolerance. J. Immunol. 171:3801–3807 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
