

Structure of lipid kinase p110β/p85β elucidates an unusual SH2-domain-mediated inhibitory mechanism
- PMID: 21362552
- PMCID: PMC3670040
- DOI: 10.1016/j.molcel.2011.01.026
Structure of lipid kinase p110β/p85β elucidates an unusual SH2-domain-mediated inhibitory mechanism
Abstract
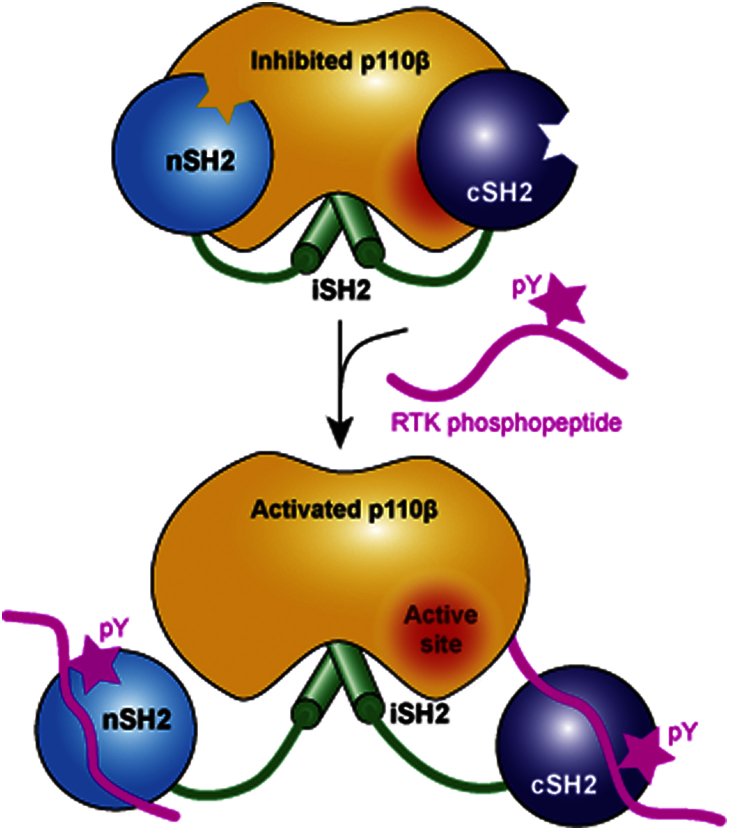
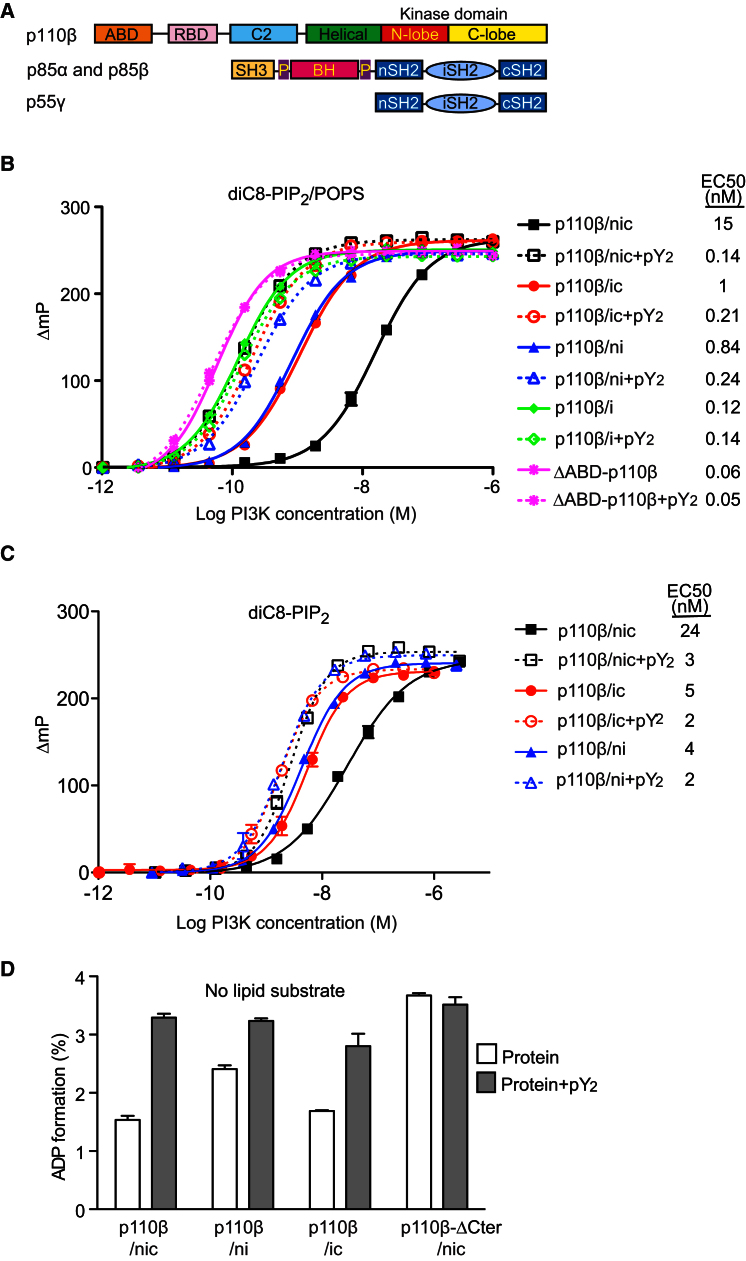
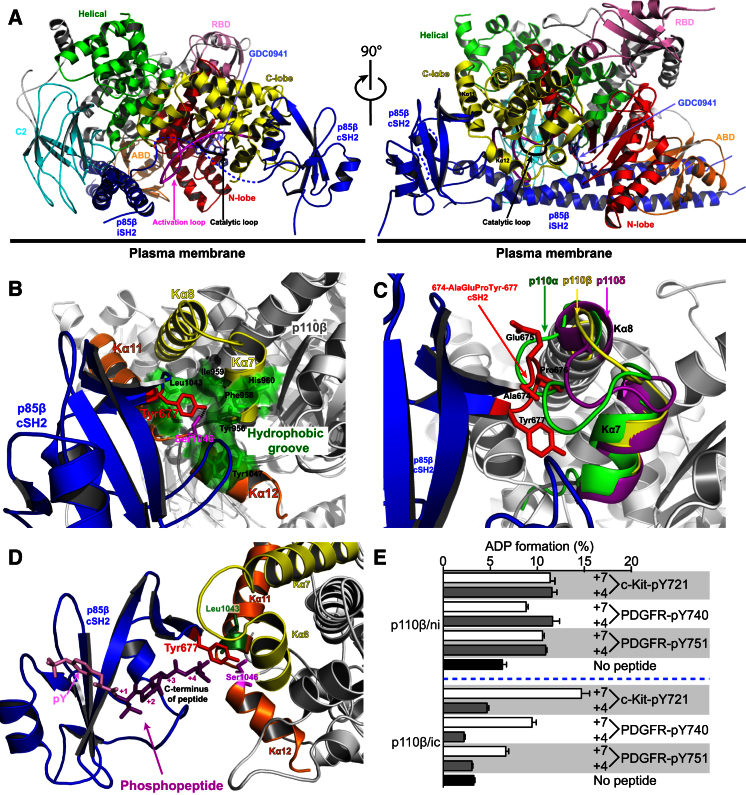
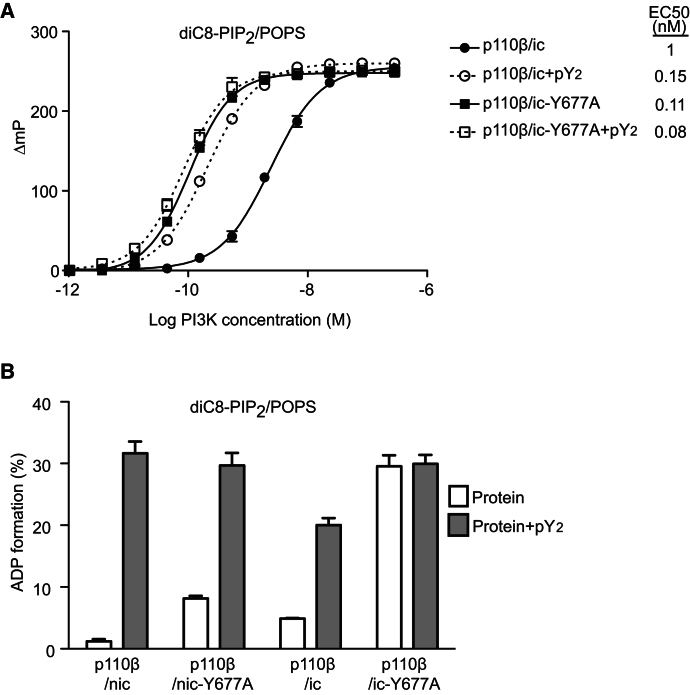
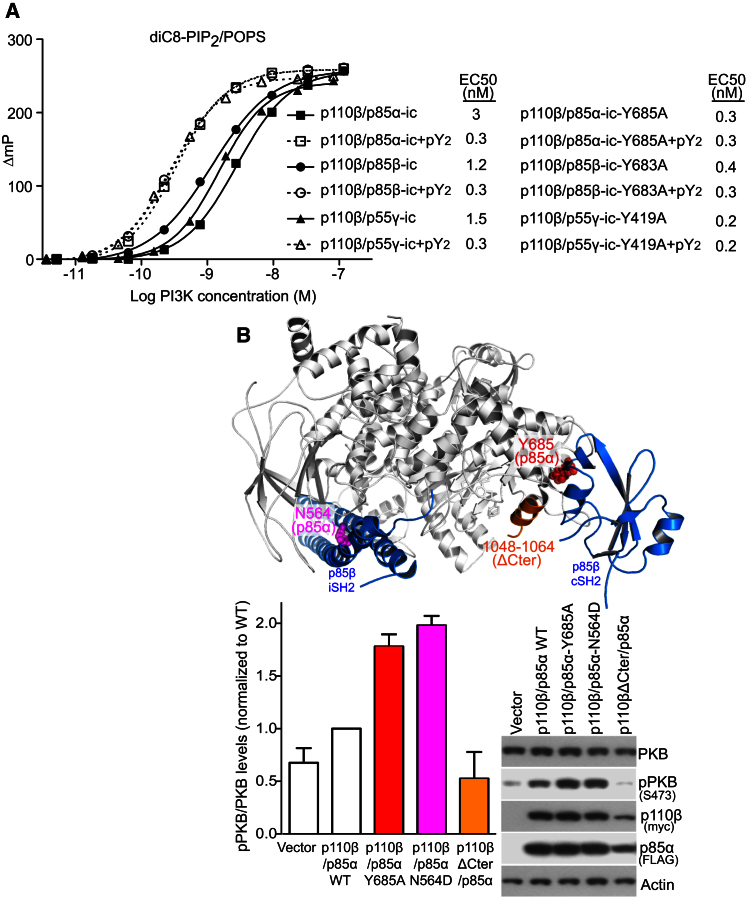
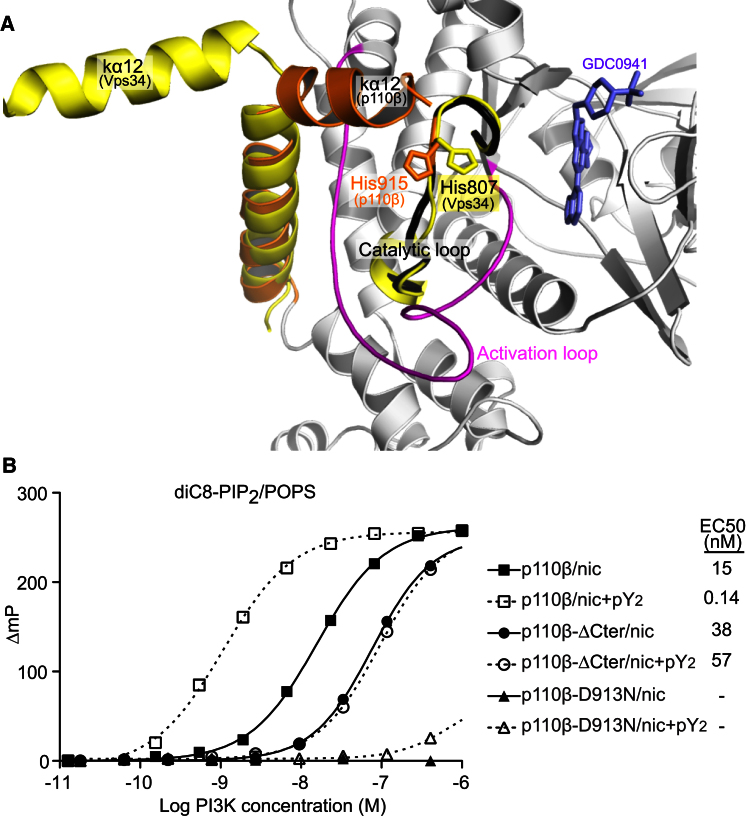
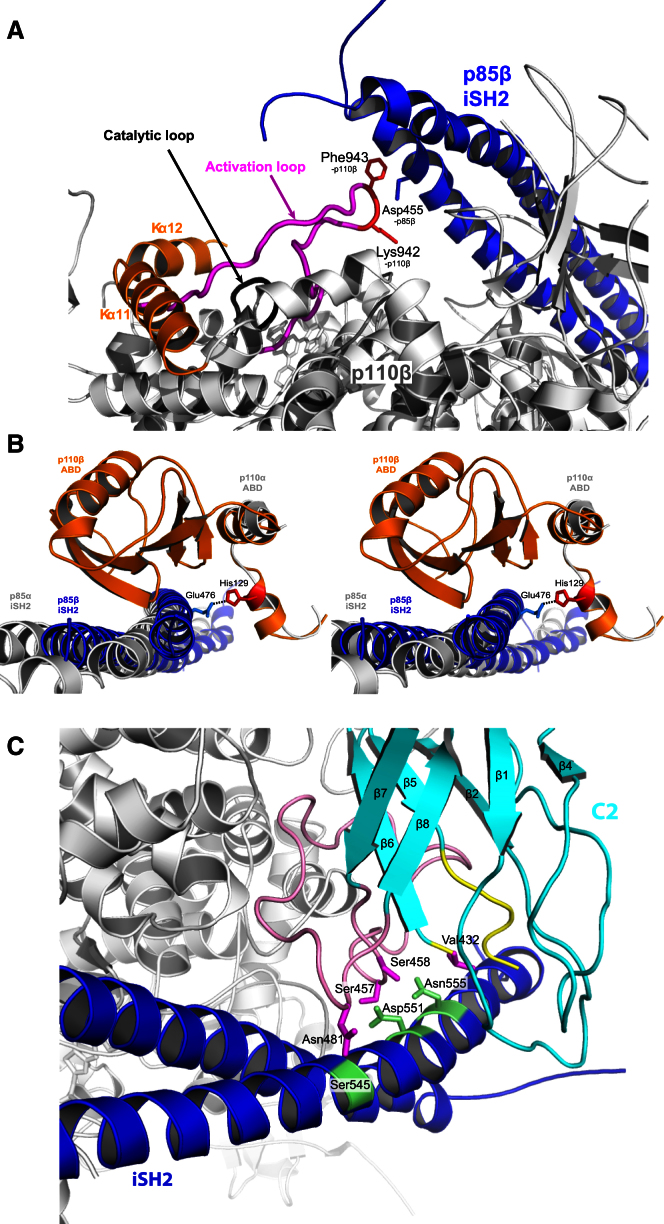
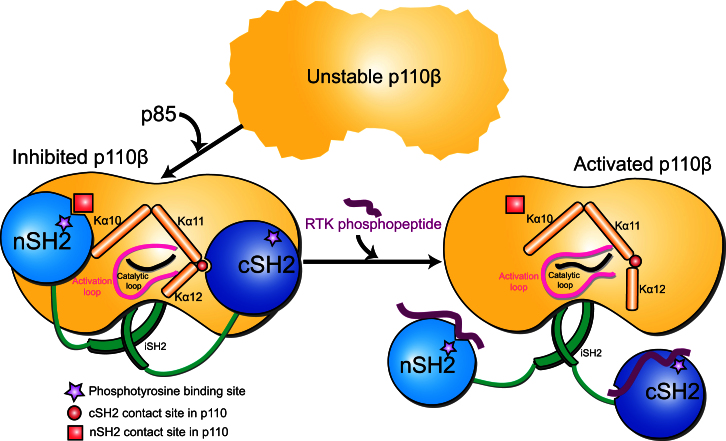
Phosphoinositide 3-kinases (PI3Ks) are essential for cell growth, migration, and survival. The structure of a p110β/p85β complex identifies an inhibitory function for the C-terminal SH2 domain (cSH2) of the p85 regulatory subunit. Mutagenesis of a cSH2 contact residue activates downstream signaling in cells. This inhibitory contact ties up the C-terminal region of the p110β catalytic subunit, which is essential for lipid kinase activity. In vitro, p110β basal activity is tightly restrained by contacts with three p85 domains: the cSH2, nSH2, and iSH2. RTK phosphopeptides relieve inhibition by nSH2 and cSH2 using completely different mechanisms. The binding site for the RTK's pYXXM motif is exposed on the cSH2, requiring an extended RTK motif to reach and disrupt the inhibitory contact with p110β. This contrasts with the nSH2 where the pY-binding site itself forms the inhibitory contact. This establishes an unusual mechanism by which p85 SH2 domains contribute to RTK signaling specificities.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
PI3K p110β: more tightly controlled or constitutively active?Mol Cell. 2011 Mar 4;41(5):499-501. doi: 10.1016/j.molcel.2011.02.017. Mol Cell. 2011. PMID: 21362546 No abstract available.
References
-
- Bondeva T., Pirola L., Bulgarelli-Leva G., Rubio I., Wetzker R., Wymann M.P. Bifurcation of lipid and protein kinase signals of PI3Kgamma to the protein kinases PKB and MAPK. Science. 1998;282:293–296. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
