

Coupled 5' nucleotide recognition and processivity in Xrn1-mediated mRNA decay
- PMID: 21362555
- PMCID: PMC3090138
- DOI: 10.1016/j.molcel.2011.02.004
Coupled 5' nucleotide recognition and processivity in Xrn1-mediated mRNA decay
Abstract
Messenger RNA decay plays a central role in the regulation and surveillance of eukaryotic gene expression. The conserved multidomain exoribonuclease Xrn1 targets cytoplasmic RNA substrates marked by a 5' monophosphate for processive 5'-to-3' degradation by an unknown mechanism. Here, we report the crystal structure of an Xrn1-substrate complex. The single-stranded substrate is held in place by stacking of the 5'-terminal trinucleotide between aromatic side chains while a highly basic pocket specifically recognizes the 5' phosphate. Mutations of residues involved in binding the 5'-terminal nucleotide impair Xrn1 processivity. The substrate recognition mechanism allows Xrn1 to couple processive hydrolysis to duplex melting in RNA substrates with sufficiently long single-stranded 5' overhangs. The Xrn1-substrate complex structure thus rationalizes the exclusive specificity of Xrn1 for 5'-monophosphorylated substrates, ensuring fidelity of mRNA turnover, and posits a model for translocation-coupled unwinding of structured RNA substrates.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
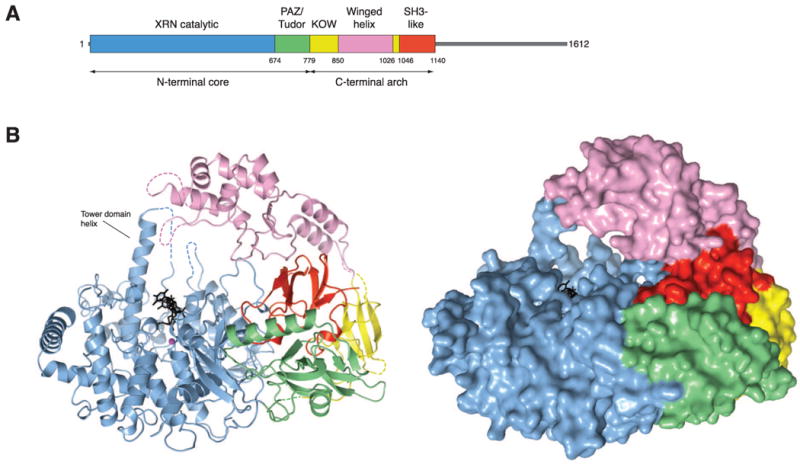
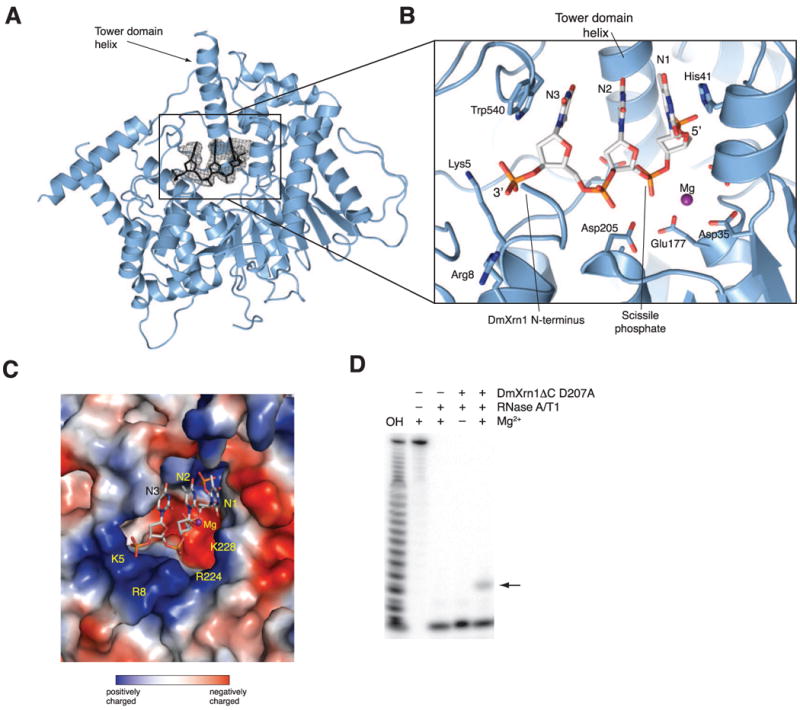
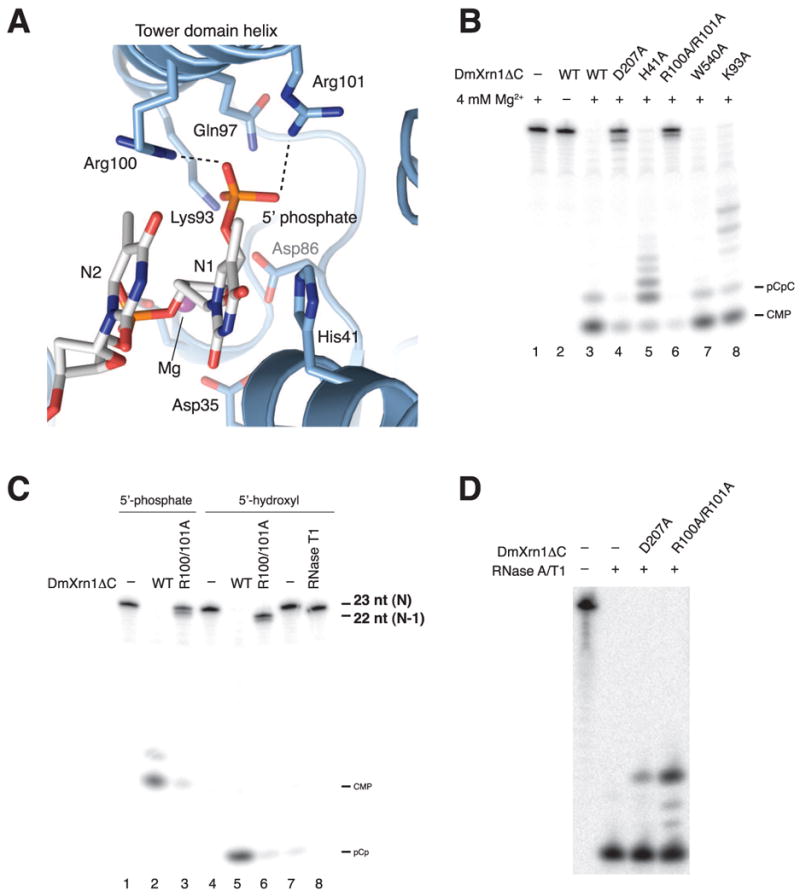
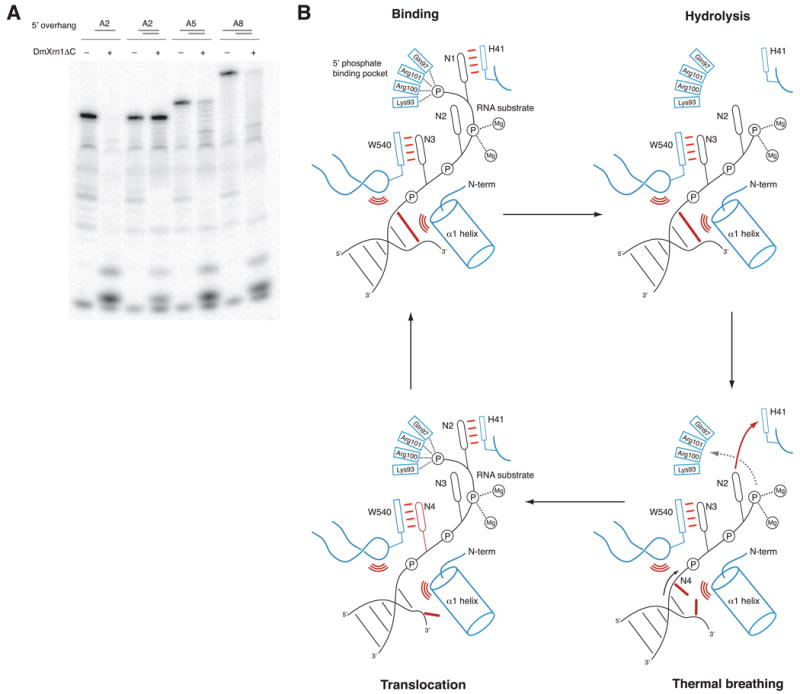
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
