

Localized delivery of interferon-β by Lactobacillus exacerbates experimental colitis
- PMID: 21365015
- PMCID: PMC3041828
- DOI: 10.1371/journal.pone.0016967
Localized delivery of interferon-β by Lactobacillus exacerbates experimental colitis
Abstract
Background: There have been conflicting reports of the role of Type I interferons (IFN) in inflammatory bowel disease (IBD). Clinical trials have shown potent efficacy of systemic interferon-beta (IFN-β) in inducing remission of ulcerative colitis. Likewise, IFNAR1(-/-) mice display an increased sensitivity to dextran sulfate sodium (DSS)-induced colitis, suggesting Type I IFN play a protective role during inflammation of the gut. Curiously, however, there have also been reports detailing the spontaneous development of IBD in patients receiving systemic IFN-β therapy for multiple sclerosis or hepatitis.
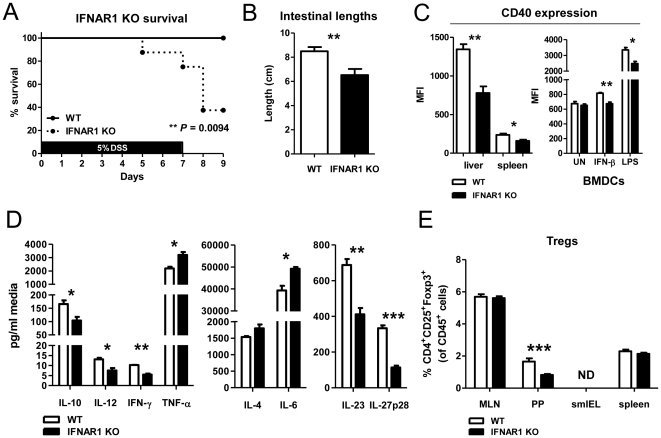
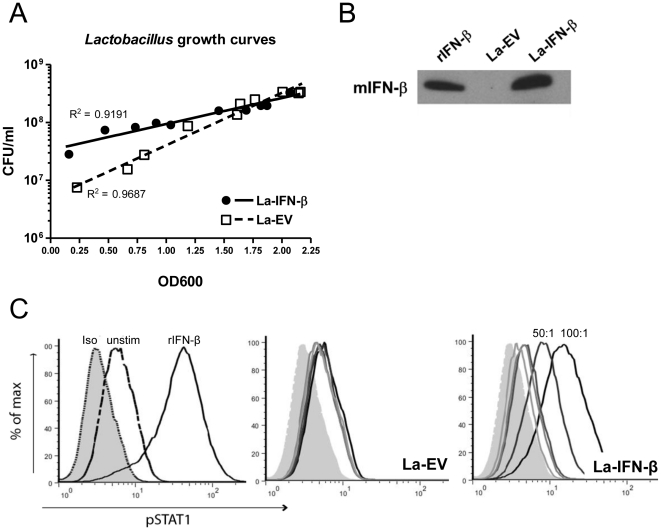
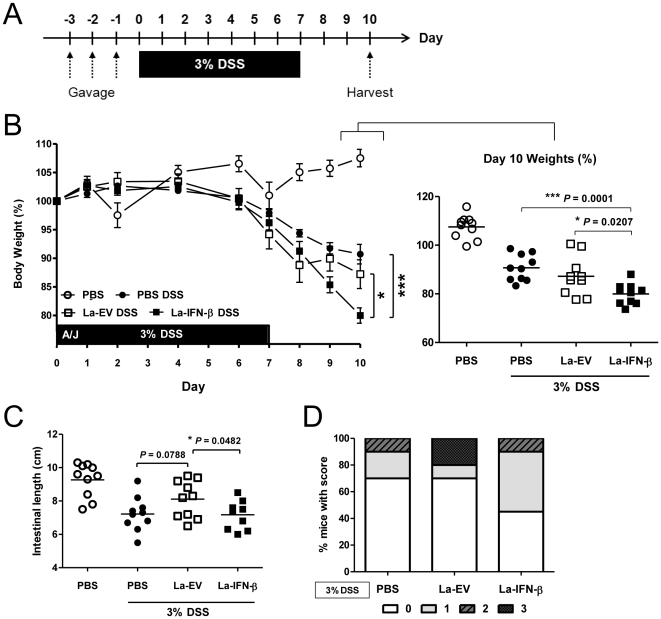
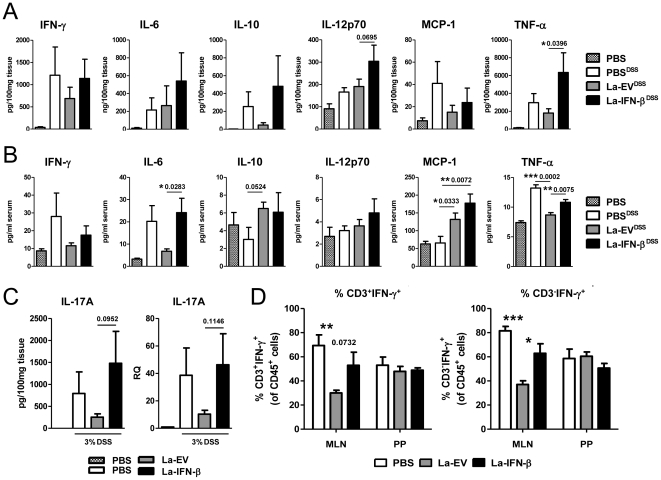
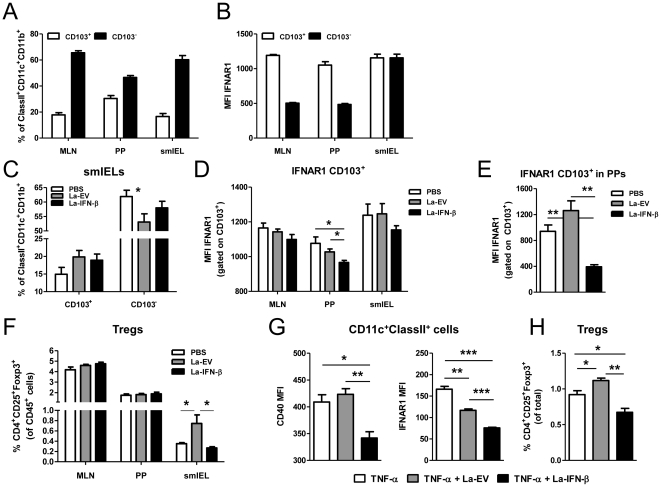
Methodology/principal findings: To investigate the effects of local administration of IFN-β on a murine model of colitis, we developed a transgenic Lactobacillus acidophilus strain that constitutively expresses IFN-β (La-IFN-β). While pretreatment of mice with control Lactobacillus (La-EV) provided slight protective benefits, La-IFN-β increased sensitivity to DSS. Analysis showed colitic mice pretreated with La-IFN-β had increased production of TNF-α, IFN-γ, IL-17A and IL-13 by intestinal tissues and decreased regulatory T cells (Tregs) in their small intestine. Examination of CD103(+) dendritic cells (DCs) in the Peyer's patches revealed that IFNAR1 expression was dramatically reduced by La-IFN-β. Similarly, bone marrow-derived DCs matured with La-IFN-β experienced a 3-fold reduction of IFNAR1 and were impaired in their ability to induce Tregs.
Conclusions/significance: Our IFNAR1 expression data identifies a correlation between the loss/downregulation of IFNAR1 on DCs and exacerbation of colitis. Our data show that Lactobacillus secreting IFN-β has an immunological effect that in our model results in the exacerbation of colitis. This study underscores that the selection of therapeutics delivered by a bacterial vehicle must take into consideration the simultaneous effects of the vehicle itself.
Conflict of interest statement
Figures
References
-
- Platanias LC. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat Rev Immunol. 2005;5:375–386. - PubMed
-
- Benveniste EN, Qin H. Type I interferons as anti-inflammatory mediators. Sci STKE 2007. 2007;416:pe70. - PubMed
-
- Triantaphyllopoulos KA, Williams RO, Tailor H, Chernajovsky Y. Amelioration of collagen-induced arthritis and suppression of interferon-gamma, interleukin-12, and tumor necrosis factor alpha production by interferon-beta gene therapy. Arthritis Rheum. 1999;42:90–99. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
