

Temperature-sensitive ipl1-2/Aurora B mutation is suppressed by mutations in TOR complex 1 via the Glc7/PP1 phosphatase
- PMID: 21368139
- PMCID: PMC3053998
- DOI: 10.1073/pnas.1014406108
Temperature-sensitive ipl1-2/Aurora B mutation is suppressed by mutations in TOR complex 1 via the Glc7/PP1 phosphatase
Abstract
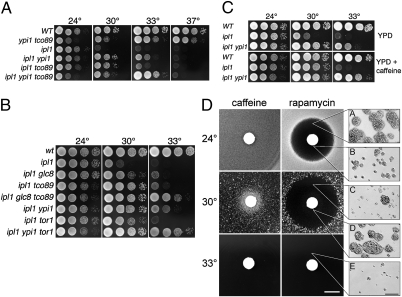
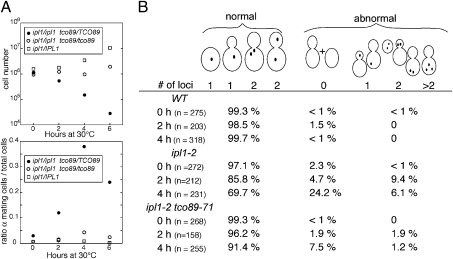
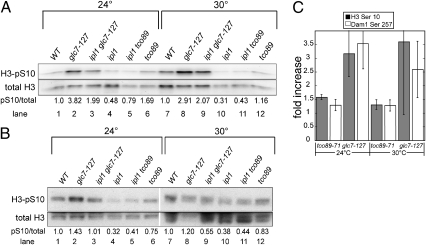
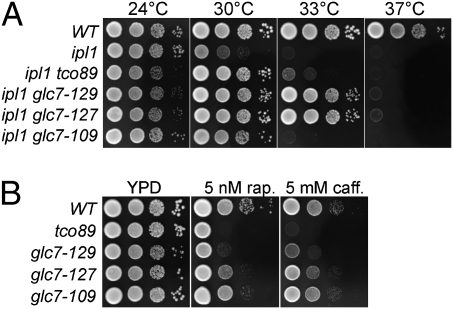
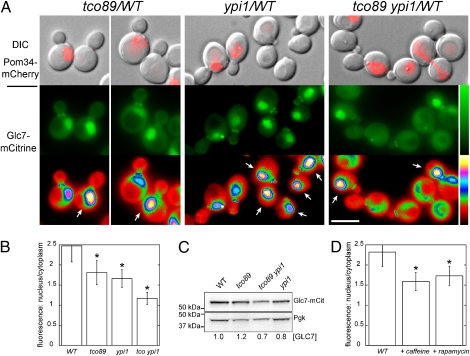
Ipl1/Aurora B is the catalytic subunit of a complex that is required for chromosome segregation and nuclear division. Before anaphase, Ipl1 localizes to kinetochores, where it is required to establish proper kinetochore-microtubule associations and regulate the spindle assembly checkpoint. The protein phosphatase Glc7/PP1 opposes Ipl1 for some of these activities. To more thoroughly characterize the Glc7 phosphatase that opposes Ipl1, we have identified mutations that suppress the thermosensitivity of an ipl1-2 mutant. In addition to mutations in genes previously associated with ipl1 suppression, we recovered a null mutant in TCO89, which encodes a subunit of the TOR complex 1 (TORC1), the conserved rapamycin-sensitive kinase activity that regulates cell growth in response to nutritional status. The temperature sensitivity of ipl1-2 can also be suppressed by null mutation of TOR1 or by administration of pharmacological TORC1 inhibitors, indicating that reduced TORC1 activity is responsible for the suppression. Suppression of the ipl1-2 growth defect is accompanied by increased fidelity of chromosome segregation and increased phosphorylation of the Ipl1 substrates histone H3 and Dam1. Nuclear Glc7 levels are reduced in a tco89 mutant, suggesting that TORC1 activity is required for the nuclear accumulation of Glc7. In addition, several mutant GLC7 alleles that suppress the temperature sensitivity of ipl1-2 exhibit negative synthetic genetic interactions with TORC1 mutants. Together, our results suggest that TORC1 positively regulates the Glc7 activity that opposes Ipl1 and provide a mechanism to tie nutritional status with mitotic regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Suppressors of ipl1-2 in components of a Glc7 phosphatase complex, Cdc48 AAA ATPase, TORC1, and the kinetochore.G3 (Bethesda). 2012 Dec;2(12):1687-701. doi: 10.1534/g3.112.003814. Epub 2012 Dec 1. G3 (Bethesda). 2012. PMID: 23275890 Free PMC article.
-
Glc7/protein phosphatase 1 regulatory subunits can oppose the Ipl1/aurora protein kinase by redistributing Glc7.Mol Cell Biol. 2006 Apr;26(7):2648-60. doi: 10.1128/MCB.26.7.2648-2660.2006. Mol Cell Biol. 2006. PMID: 16537909 Free PMC article.
-
The budding yeast Cdc48(Shp1) complex promotes cell cycle progression by positive regulation of protein phosphatase 1 (Glc7).PLoS One. 2013;8(2):e56486. doi: 10.1371/journal.pone.0056486. Epub 2013 Feb 13. PLoS One. 2013. PMID: 23418575 Free PMC article.
-
Protein kinases in mitotic phosphorylation of budding yeast CENP-A.Curr Genet. 2019 Dec;65(6):1325-1332. doi: 10.1007/s00294-019-00997-5. Epub 2019 May 22. Curr Genet. 2019. PMID: 31119371 Review.
-
Function of protein phosphatase-1, Glc7, in Saccharomyces cerevisiae.Adv Appl Microbiol. 2010;73:27-59. doi: 10.1016/S0065-2164(10)73002-1. Adv Appl Microbiol. 2010. PMID: 20800758 Review.
Cited by
-
Phosphoregulation of the yeast Pma1 H+-ATPase autoinhibitory domain involves the Ptk1/2 kinases and the Glc7 PP1 phosphatase and is under TORC1 control.PLoS Genet. 2024 Jan 16;20(1):e1011121. doi: 10.1371/journal.pgen.1011121. eCollection 2024 Jan. PLoS Genet. 2024. PMID: 38227612 Free PMC article.
-
The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment.Int J Mol Sci. 2019 Dec 7;20(24):6182. doi: 10.3390/ijms20246182. Int J Mol Sci. 2019. PMID: 31817904 Free PMC article. Review.
-
Target of rapamycin signaling regulates high mobility group protein association to chromatin, which functions to suppress necrotic cell death.Epigenetics Chromatin. 2013 Sep 2;6(1):29. doi: 10.1186/1756-8935-6-29. Epigenetics Chromatin. 2013. PMID: 24044743 Free PMC article.
-
Interactions between the kinetochore complex and the protein kinase A pathway in Saccharomyces cerevisiae.G3 (Bethesda). 2012 Jul;2(7):831-41. doi: 10.1534/g3.112.002675. Epub 2012 Jul 1. G3 (Bethesda). 2012. PMID: 22870406 Free PMC article.
-
Chemical suppression of defects in mitotic spindle assembly, redox control, and sterol biosynthesis by hydroxyurea.G3 (Bethesda). 2014 Jan 10;4(1):39-48. doi: 10.1534/g3.113.009100. G3 (Bethesda). 2014. PMID: 24192836 Free PMC article.
References
-
- Ruchaud S, Carmena M, Earnshaw WC. Chromosomal passengers: Conducting cell division. Nat Rev Mol Cell Biol. 2007;8:798–812. - PubMed
-
- He X, Rines DR, Espelin CW, Sorger PK. Molecular analysis of kinetochore-microtubule attachment in budding yeast. Cell. 2001;106:195–206. - PubMed
-
- Tanaka TU, et al. Evidence that the Ipl1-Sli15 (Aurora kinase-INCENP) complex promotes chromosome bi-orientation by altering kinetochore-spindle pole connections. Cell. 2002;108:317–329. - PubMed
-
- Lampson MA, Renduchitala K, Khodjakov A, Kapoor TM. Correcting improper chromosome-spindle attachments during cell division. Nat Cell Biol. 2004;6:232–237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
