

Spatial effects on species persistence and implications for biodiversity
- PMID: 21368181
- PMCID: PMC3060266
- DOI: 10.1073/pnas.1017274108
Spatial effects on species persistence and implications for biodiversity
Abstract
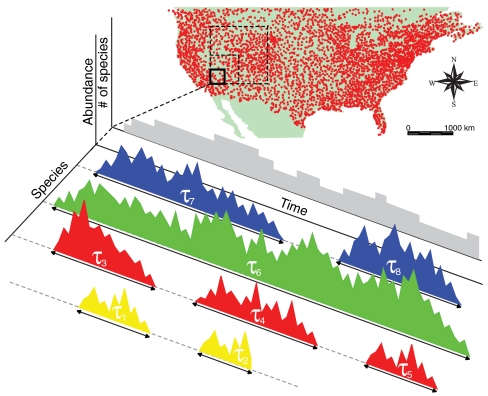
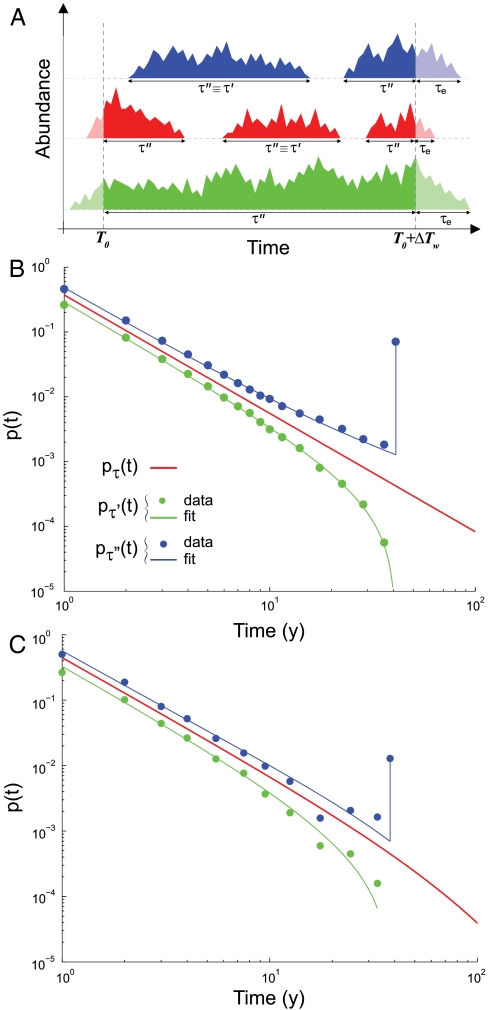
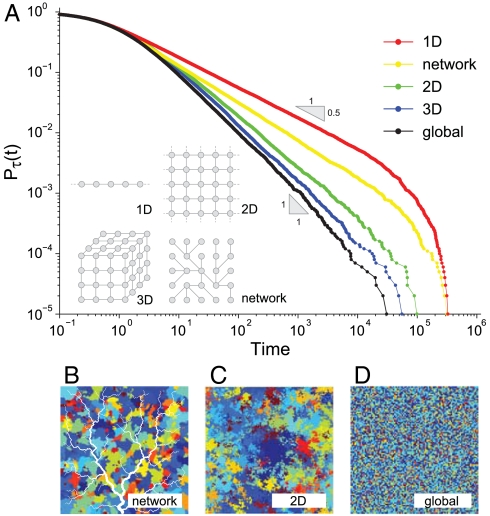
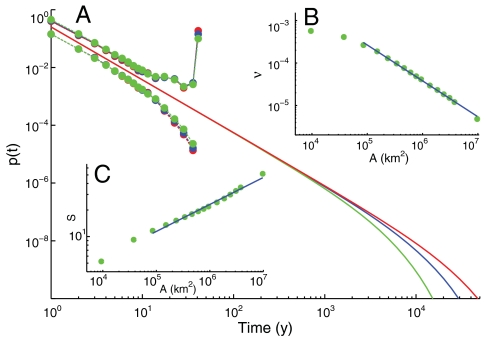
Natural ecosystems are characterized by striking diversity of form and functions and yet exhibit deep symmetries emerging across scales of space, time, and organizational complexity. Species-area relationships and species-abundance distributions are examples of emerging patterns irrespective of the details of the underlying ecosystem functions. Here we present empirical and theoretical evidence for a new macroecological pattern related to the distributions of local species persistence times, defined as the time spans between local colonizations and extinctions in a given geographic region. Empirical distributions pertaining to two different taxa, breeding birds and herbaceous plants, analyzed in a framework that accounts for the finiteness of the observational period exhibit power-law scaling limited by a cutoff determined by the rate of emergence of new species. In spite of the differences between taxa and spatial scales of analysis, the scaling exponents are statistically indistinguishable from each other and significantly different from those predicted by existing models. We theoretically investigate how the scaling features depend on the structure of the spatial interaction network and show that the empirical scaling exponents are reproduced once a two-dimensional isotropic texture is used, regardless of the details of the ecological interactions. The framework developed here also allows to link the cutoff time scale with the spatial scale of analysis, and the persistence-time distribution to the species-area relationship. We conclude that the inherent coherence obtained between spatial and temporal macroecological patterns points at a seemingly general feature of the dynamical evolution of ecosystems.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
On species persistence-time distributions.J Theor Biol. 2012 Jun 21;303:15-24. doi: 10.1016/j.jtbi.2012.02.022. Epub 2012 Mar 3. J Theor Biol. 2012. PMID: 22763130
-
Spatial scale, abundance and the species-energy relationship in British birds.J Anim Ecol. 2008 Mar;77(2):395-405. doi: 10.1111/j.1365-2656.2007.01332.x. Epub 2007 Nov 13. J Anim Ecol. 2008. PMID: 18005031
-
Emergent dual scaling of riverine biodiversity.Proc Natl Acad Sci U S A. 2021 Nov 23;118(47):e2105574118. doi: 10.1073/pnas.2105574118. Proc Natl Acad Sci U S A. 2021. PMID: 34795054 Free PMC article.
-
The spatial scaling of species interaction networks.Nat Ecol Evol. 2018 May;2(5):782-790. doi: 10.1038/s41559-018-0517-3. Epub 2018 Apr 16. Nat Ecol Evol. 2018. PMID: 29662224 Review.
-
Linking biodiversity patterns by autocorrelated random sampling.Am J Bot. 2011 Mar;98(3):481-502. doi: 10.3732/ajb.1000509. Epub 2011 Mar 2. Am J Bot. 2011. PMID: 21613141 Review.
Cited by
-
Coexistence of species with different dispersal across landscapes: a critical role of spatial correlation in disturbance.Proc Biol Sci. 2016 May 11;283(1830):20160537. doi: 10.1098/rspb.2016.0537. Proc Biol Sci. 2016. PMID: 27147101 Free PMC article.
-
Microbial coexistence through chemical-mediated interactions.Nat Commun. 2019 May 3;10(1):2052. doi: 10.1038/s41467-019-10062-x. Nat Commun. 2019. PMID: 31053707 Free PMC article.
-
Macroecological dynamics of gut microbiota.Nat Microbiol. 2020 May;5(5):768-775. doi: 10.1038/s41564-020-0685-1. Epub 2020 Apr 13. Nat Microbiol. 2020. PMID: 32284567
-
Forecasting unprecedented ecological fluctuations.PLoS Comput Biol. 2020 Jun 29;16(6):e1008021. doi: 10.1371/journal.pcbi.1008021. eCollection 2020 Jun. PLoS Comput Biol. 2020. PMID: 32598364 Free PMC article.
-
Species extinction thresholds in the face of spatially correlated periodic disturbance.Sci Rep. 2015 Oct 20;5:15455. doi: 10.1038/srep15455. Sci Rep. 2015. PMID: 26482293 Free PMC article.
References
-
- Diamond J. The present, past and future of human-caused exctinctions. Philos Trans R Soc London B. 1989;325:469–477. - PubMed
-
- Brown JH. Macroecology. Chicago: Univ Chicago Press; 1995.
-
- Thomas C, et al. Extinction risk from climate change. Nature. 2004;427:145–148. - PubMed
-
- Svenning J.-C, Condit R. Biodiversity in a warmer world. Science. 2008;322:206–207. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
