

Niche of harmful alga Aureococcus anophagefferens revealed through ecogenomics
- PMID: 21368207
- PMCID: PMC3060233
- DOI: 10.1073/pnas.1016106108
Niche of harmful alga Aureococcus anophagefferens revealed through ecogenomics
Abstract
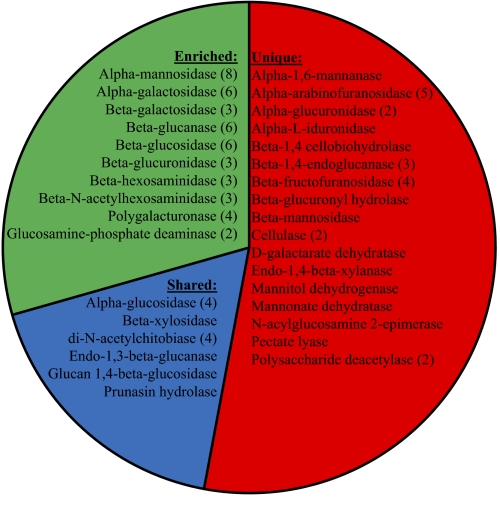
Harmful algal blooms (HABs) cause significant economic and ecological damage worldwide. Despite considerable efforts, a comprehensive understanding of the factors that promote these blooms has been lacking, because the biochemical pathways that facilitate their dominance relative to other phytoplankton within specific environments have not been identified. Here, biogeochemical measurements showed that the harmful alga Aureococcus anophagefferens outcompeted co-occurring phytoplankton in estuaries with elevated levels of dissolved organic matter and turbidity and low levels of dissolved inorganic nitrogen. We subsequently sequenced the genome of A. anophagefferens and compared its gene complement with those of six competing phytoplankton species identified through metaproteomics. Using an ecogenomic approach, we specifically focused on gene sets that may facilitate dominance within the environmental conditions present during blooms. A. anophagefferens possesses a larger genome (56 Mbp) and has more genes involved in light harvesting, organic carbon and nitrogen use, and encoding selenium- and metal-requiring enzymes than competing phytoplankton. Genes for the synthesis of microbial deterrents likely permit the proliferation of this species, with reduced mortality losses during blooms. Collectively, these findings suggest that anthropogenic activities resulting in elevated levels of turbidity, organic matter, and metals have opened a niche within coastal ecosystems that ideally suits the unique genetic capacity of A. anophagefferens and thus, has facilitated the proliferation of this and potentially other HABs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Sunda WG, Graneli E, Gobler CJ. Positive feedback and the development and persistence of ecosystem disruptive algal blooms. J Phycol. 2006;42:963–974.
-
- Smayda TJ. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol Oceanogr. 1997;42:1137–1153.
-
- Hoagland P, Scatasta S. In: Ecology of Harmful Algae. Graneli E, Turner J, editors. Berlin: Springer; 2006. pp. 391–402.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
