

Plant centromeric retrotransposons: a structural and cytogenetic perspective
- PMID: 21371312
- PMCID: PMC3059260
- DOI: 10.1186/1759-8753-2-4
Plant centromeric retrotransposons: a structural and cytogenetic perspective
Abstract
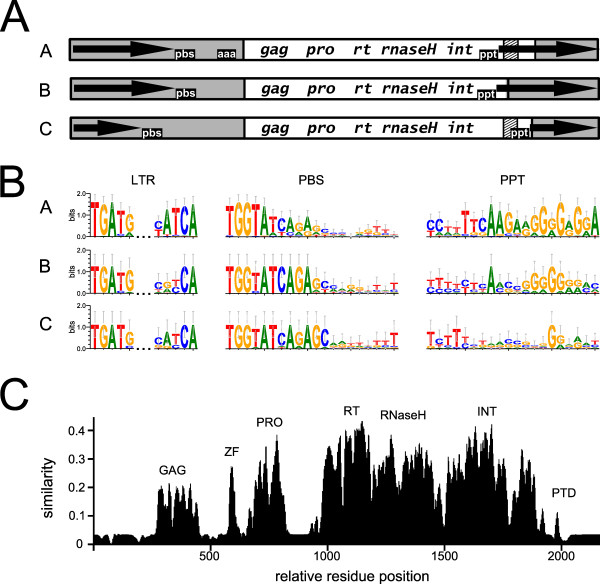
Background: The centromeric and pericentromeric regions of plant chromosomes are colonized by Ty3/gypsy retrotransposons, which, on the basis of their reverse transcriptase sequences, form the chromovirus CRM clade. Despite their potential importance for centromere evolution and function, they have remained poorly characterized. In this work, we aimed to carry out a comprehensive survey of CRM clade elements with an emphasis on their diversity, structure, chromosomal distribution and transcriptional activity.
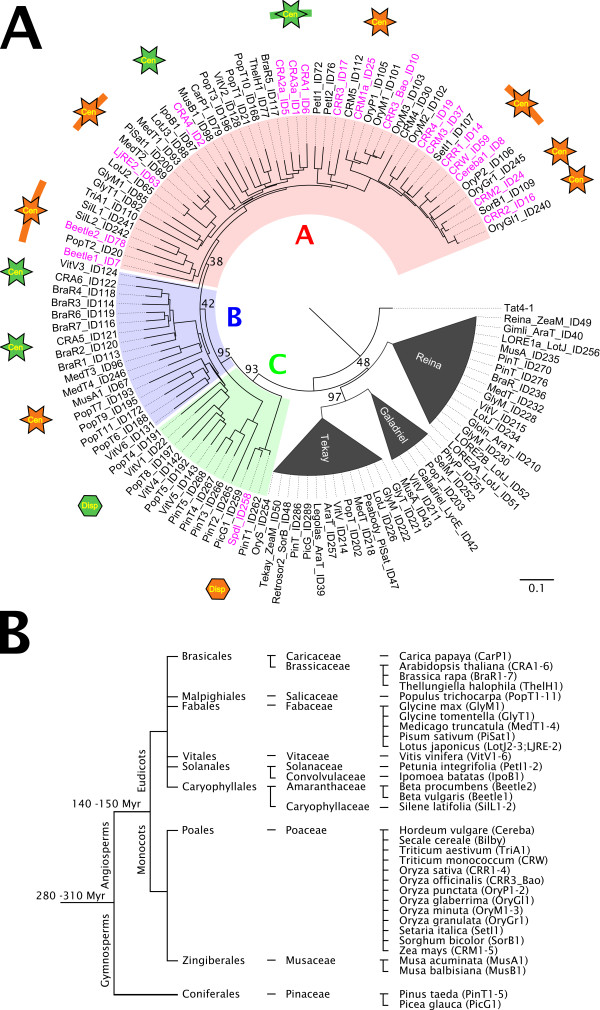
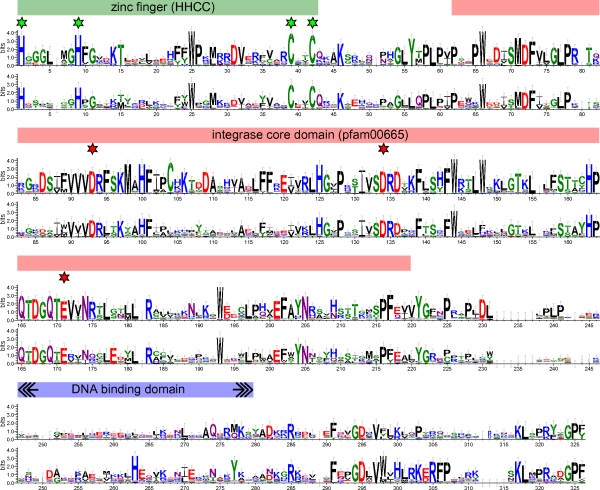
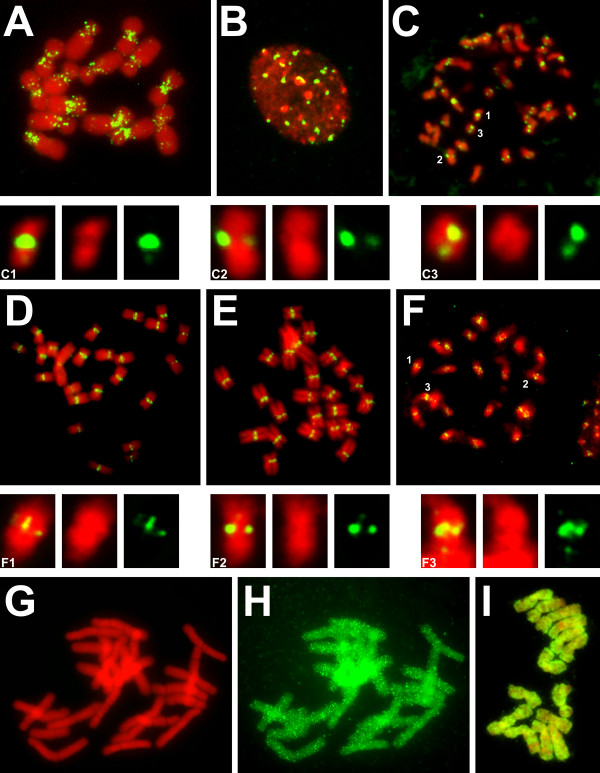
Results: We have surveyed a set of 190 CRM elements belonging to 81 different retrotransposon families, derived from 33 host species and falling into 12 plant families. The sequences at the C-terminus of their integrases were unexpectedly heterogeneous, despite the understanding that they are responsible for targeting to the centromere. This variation allowed the division of the CRM clade into the three groups A, B and C, and the members of each differed considerably with respect to their chromosomal distribution. The differences in chromosomal distribution coincided with variation in the integrase C-terminus sequences possessing a putative targeting domain (PTD). A majority of the group A elements possess the CR motif and are concentrated in the centromeric region, while members of group C have the type II chromodomain and are dispersed throughout the genome. Although representatives of the group B lack a PTD of any type, they appeared to be localized preferentially in the centromeres of tested species. All tested elements were found to be transcriptionally active.
Conclusions: Comprehensive analysis of the CRM clade elements showed that genuinely centromeric retrotransposons represent only a fraction of the CRM clade (group A). These centromeric retrotransposons represent an active component of centromeres of a wide range of angiosperm species, implying that they play an important role in plant centromere evolution. In addition, their transcriptional activity is consistent with the notion that the transcription of centromeric retrotransposons has a role in normal centromere function.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
