

JAK-STAT3 pathway regulates spinal astrocyte proliferation and neuropathic pain maintenance in rats
- PMID: 21371995
- PMCID: PMC4571138
- DOI: 10.1093/brain/awr025
JAK-STAT3 pathway regulates spinal astrocyte proliferation and neuropathic pain maintenance in rats
Abstract
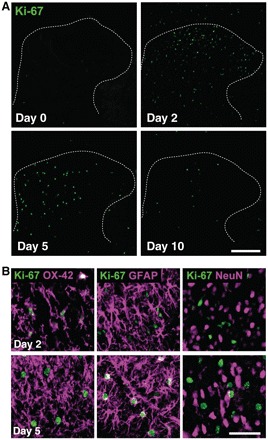
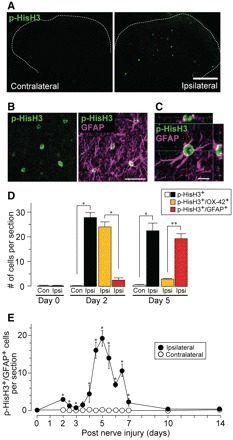
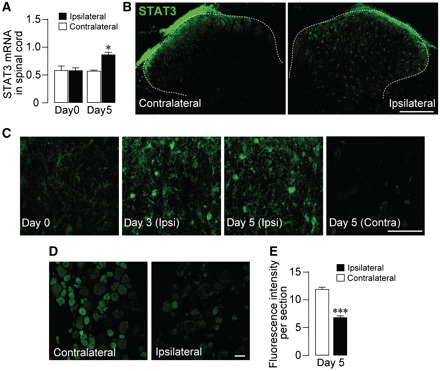
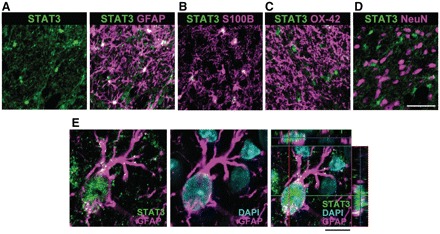
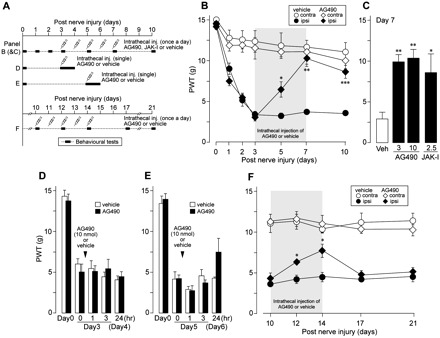
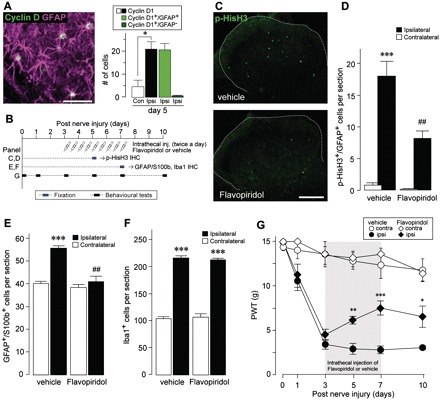
Neuropathic pain, a debilitating pain condition, is a common consequence of damage to the nervous system. Optimal treatment of neuropathic pain is a major clinical challenge because the underlying mechanisms remain unclear and currently available treatments are frequently ineffective. Emerging lines of evidence indicate that peripheral nerve injury converts resting spinal cord glia into reactive cells that are required for the development and maintenance of neuropathic pain. However, the mechanisms underlying reactive astrogliosis after nerve injury are largely unknown. In the present study, we investigated cell proliferation, a critical process in reactive astrogliosis, and determined the temporally restricted proliferation of dorsal horn astrocytes in rats with spinal nerve injury, a well-known model of neuropathic pain. We found that nerve injury-induced astrocyte proliferation requires the Janus kinase-signal transducers and activators of transcription 3 signalling pathway. Nerve injury induced a marked signal transducers and activators of transcription 3 nuclear translocation, a primary index of signal transducers and activators of transcription 3 activation, in dorsal horn astrocytes. Intrathecally administering inhibitors of Janus kinase-signal transducers and activators of transcription 3 signalling to rats with nerve injury reduced the number of proliferating dorsal horn astrocytes and produced a recovery from established tactile allodynia, a cardinal symptom of neuropathic pain that is characterized by pain hypersensitivity evoked by innocuous stimuli. Moreover, recovery from tactile allodynia was also produced by direct suppression of dividing astrocytes by intrathecal administration of the cell cycle inhibitor flavopiridol to nerve-injured rats. Together, these results imply that the Janus kinase-signal transducers and activators of transcription 3 signalling pathway are critical transducers of astrocyte proliferation and maintenance of tactile allodynia and may be a therapeutic target for neuropathic pain.
Figures

References
-
- Baron R. Mechanisms of disease: neuropathic pain–a clinical perspective. Nat Clin Pract Neurol. 2006;2:95–106. - PubMed
-
- Bonni A, Sun Y, Nadal-Vicens M, Bhatt A, Frank DA, Rozovsky I, et al. Regulation of gliogenesis in the central nervous system by the JAK-STAT signaling pathway. Science. 1997;278:477–83. - PubMed
-
- Byrnes KR, Stoica BA, Fricke S, Di Giovanni S, Faden AI. Cell cycle activation contributes to post-mitotic cell death and secondary damage after spinal cord injury. Brain. 2007;130:2977–92. - PubMed
-
- Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53:55–63. - PubMed
-
- Choi JS, Kim SY, Cha JH, Choi YS, Sung KW, Oh ST, et al. Upregulation of gp130 and STAT3 activation in the rat hippocampus following transient forebrain ischemia. Glia. 2003;41:237–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
