

DsbA2 (27 kDa Com1-like protein) of Legionella pneumophila catalyses extracytoplasmic disulphide-bond formation in proteins including the Dot/Icm type IV secretion system
- PMID: 21375592
- PMCID: PMC3086132
- DOI: 10.1111/j.1365-2958.2011.07615.x
DsbA2 (27 kDa Com1-like protein) of Legionella pneumophila catalyses extracytoplasmic disulphide-bond formation in proteins including the Dot/Icm type IV secretion system
Abstract
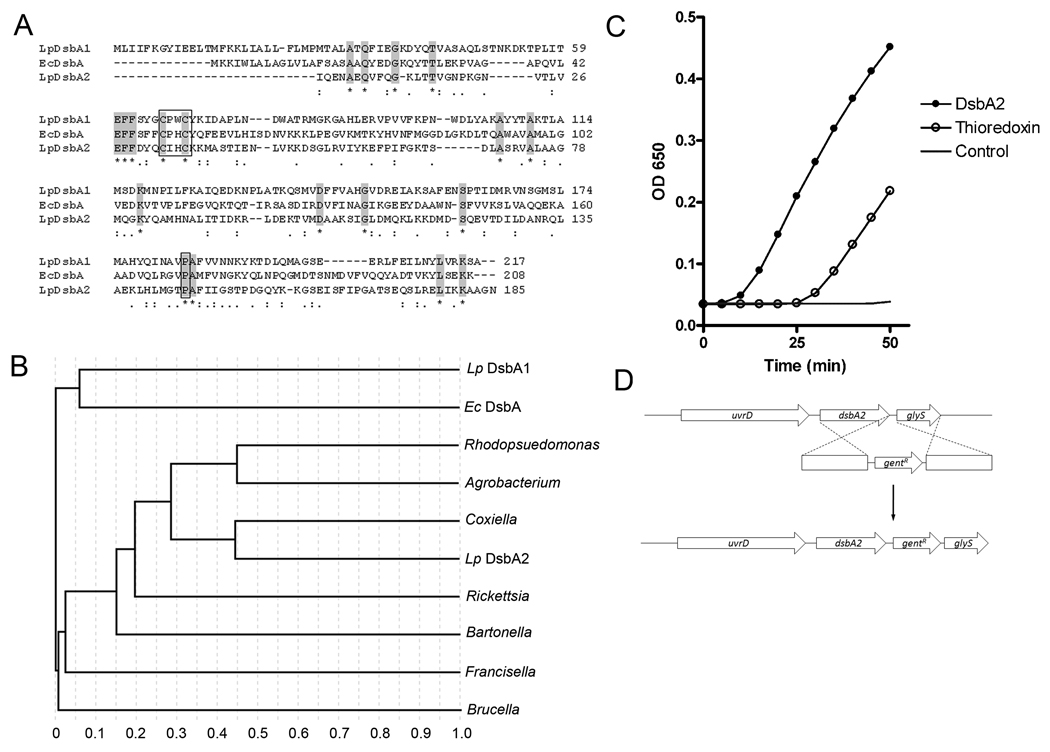
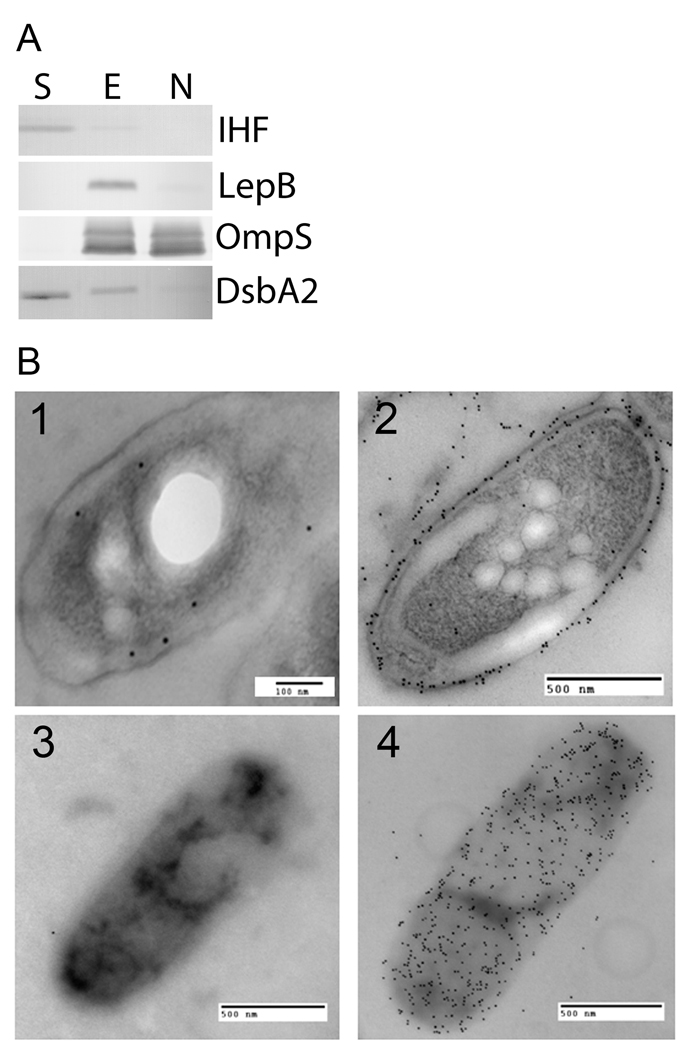
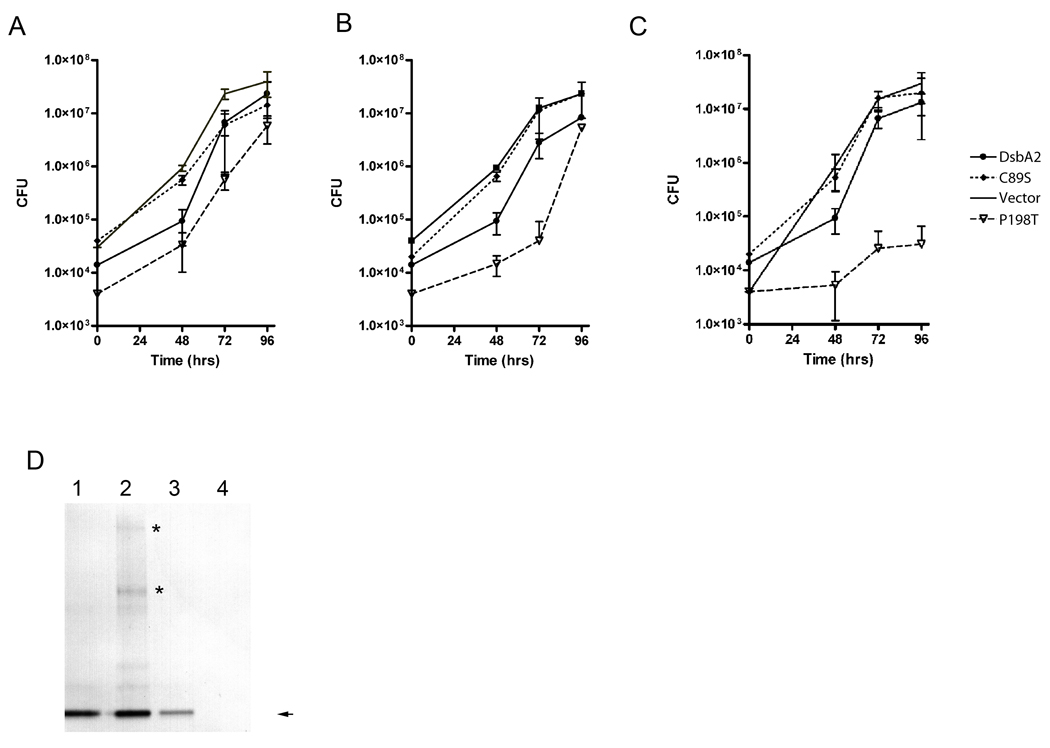
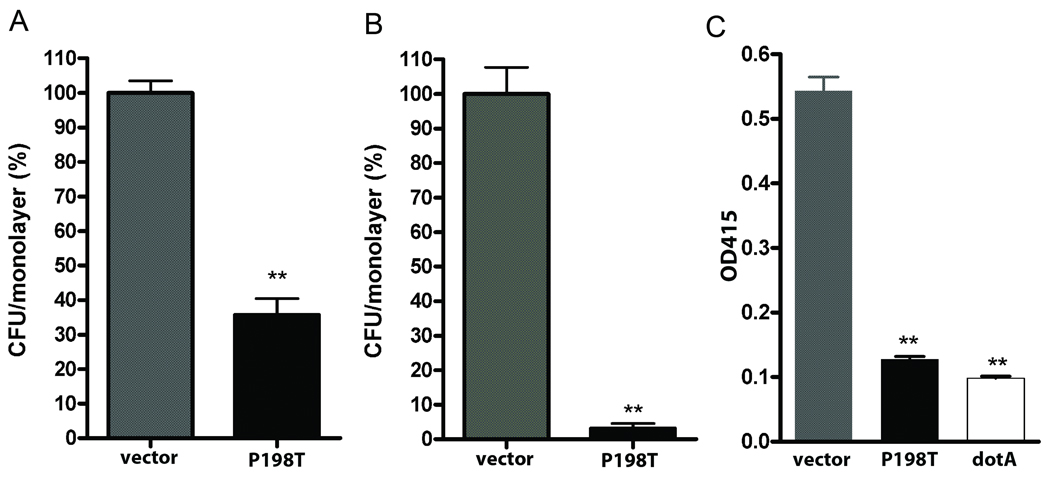
In Gram-negative bacteria, thiol oxidoreductases catalyse the formation of disulphide bonds (DSB) in extracytoplasmic proteins. In this study, we sought to identify DSB-forming proteins required for assembly of macromolecular structures in Legionella pneumophila. Here we describe two DSB-forming proteins, one annotated as dsbA1 and the other annotated as a 27 kDa outer membrane protein similar to Com1 of Coxiella burnetii, which we designate as dsbA2. Both proteins are predicted to be periplasmic, and while dsbA1 mutants were readily isolated and without phenotype, dsbA2 mutants were not obtained. To advance studies of DsbA2, a cis-proline residue at position 198 was replaced with threonine that enables formation of stable disulphide-bond complexes with substrate proteins. Expression of DsbA2 P198T mutant protein from an inducible promoter produced dominant-negative effects on DsbA2 function that resulted in loss of infectivity for amoeba and HeLa cells and loss of Dot/Icm T4SS-mediated contact haemolysis of erythrocytes. Analysis of captured DsbA2 P198T-substrate complexes from L. pneumophila by mass spectrometry identified periplasmic and outer membrane proteins that included components of the Dot/Icm T4SS. More broadly, our studies establish a DSB oxidoreductase function for the Com1 lineage of DsbA2-like proteins which appear to be conserved among those bacteria also expressing T4SS.
© 2011 Blackwell Publishing Ltd.
Figures
References
-
- Bardwell JC, McGovern K, Beckwith J. Identification of a protein required for disulfide bond formation in vivo. Cell. 1991;67:581–589. - PubMed
-
- Berger KH, Isberg RR. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol Microbiol. 1993;7:7–19. - PubMed
-
- Berk SG, Faulkner G, Garduno E, Joy MC, Ortiz-Jimenez MA, Garduno RA. Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a Dot/Icm-mediated survival mechanism. Appl Environ Microbiol. 2008;74:2187–2199. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
