

The functional role of the T1R family of receptors in sweet taste and feeding
- PMID: 21376068
- PMCID: PMC3186843
- DOI: 10.1016/j.physbeh.2011.02.030
The functional role of the T1R family of receptors in sweet taste and feeding
Abstract
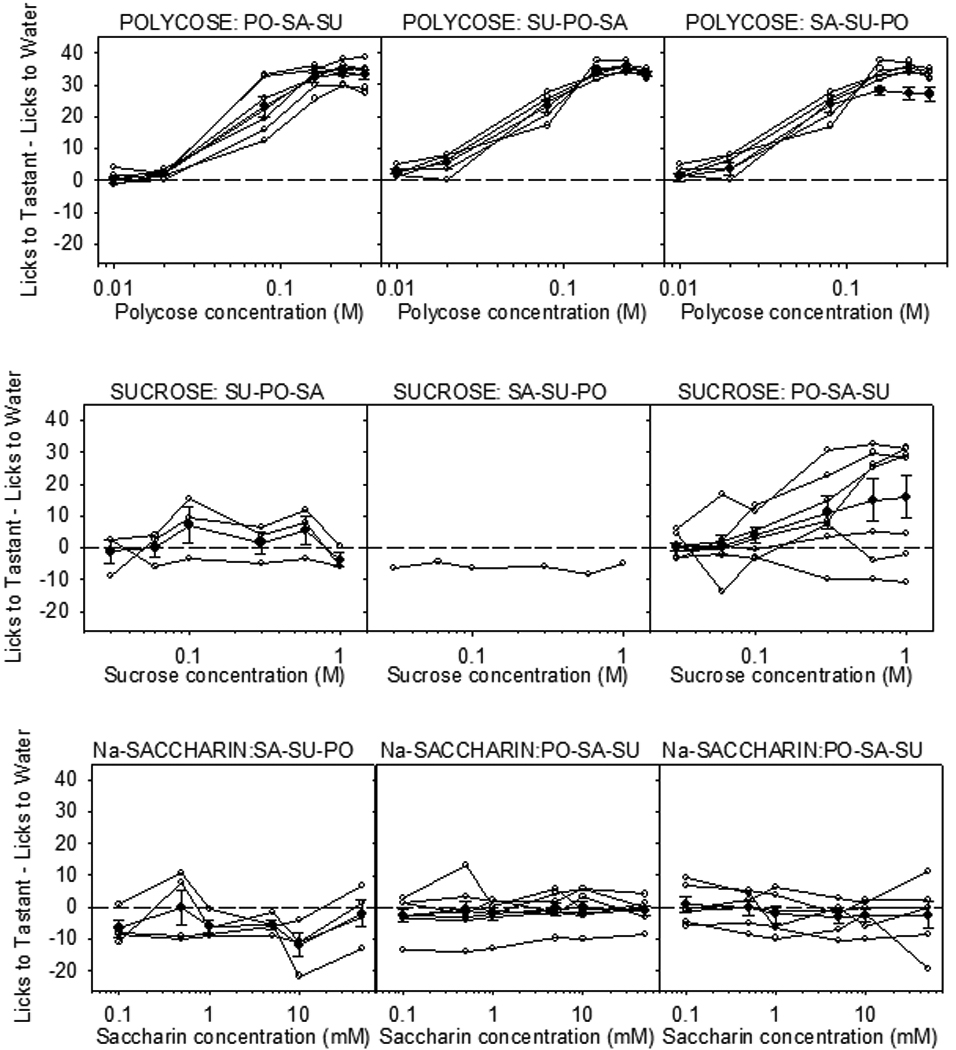
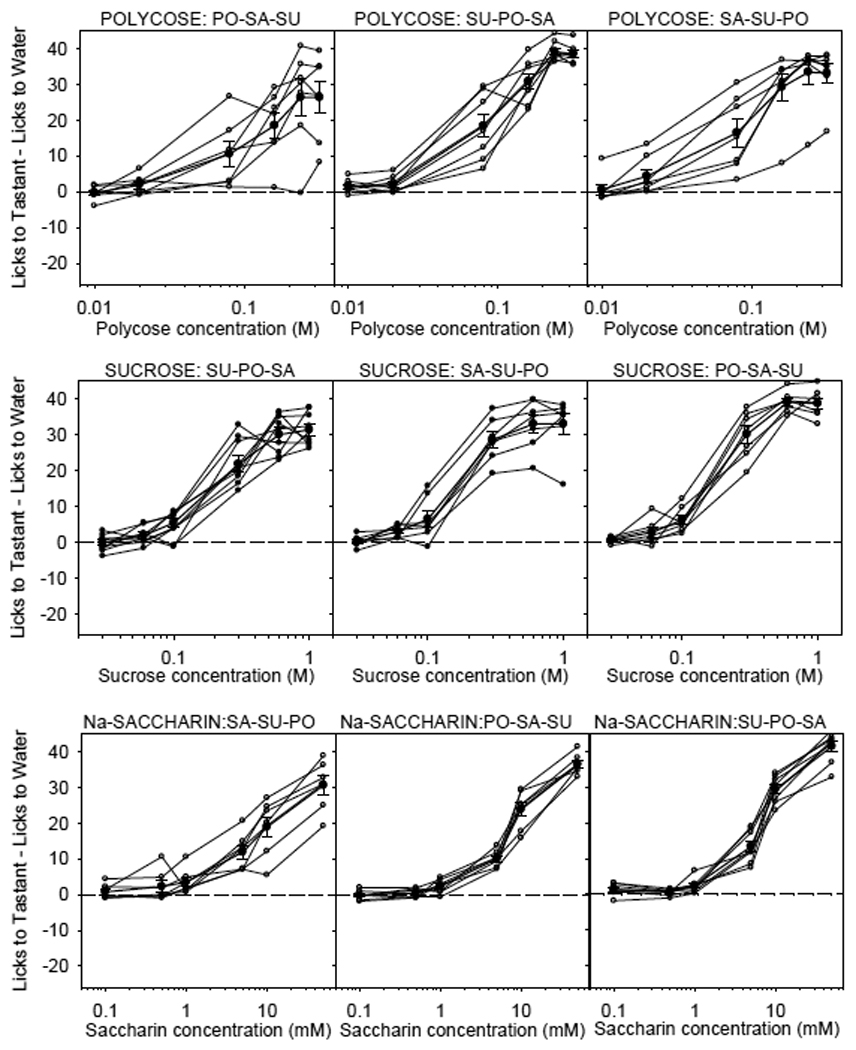
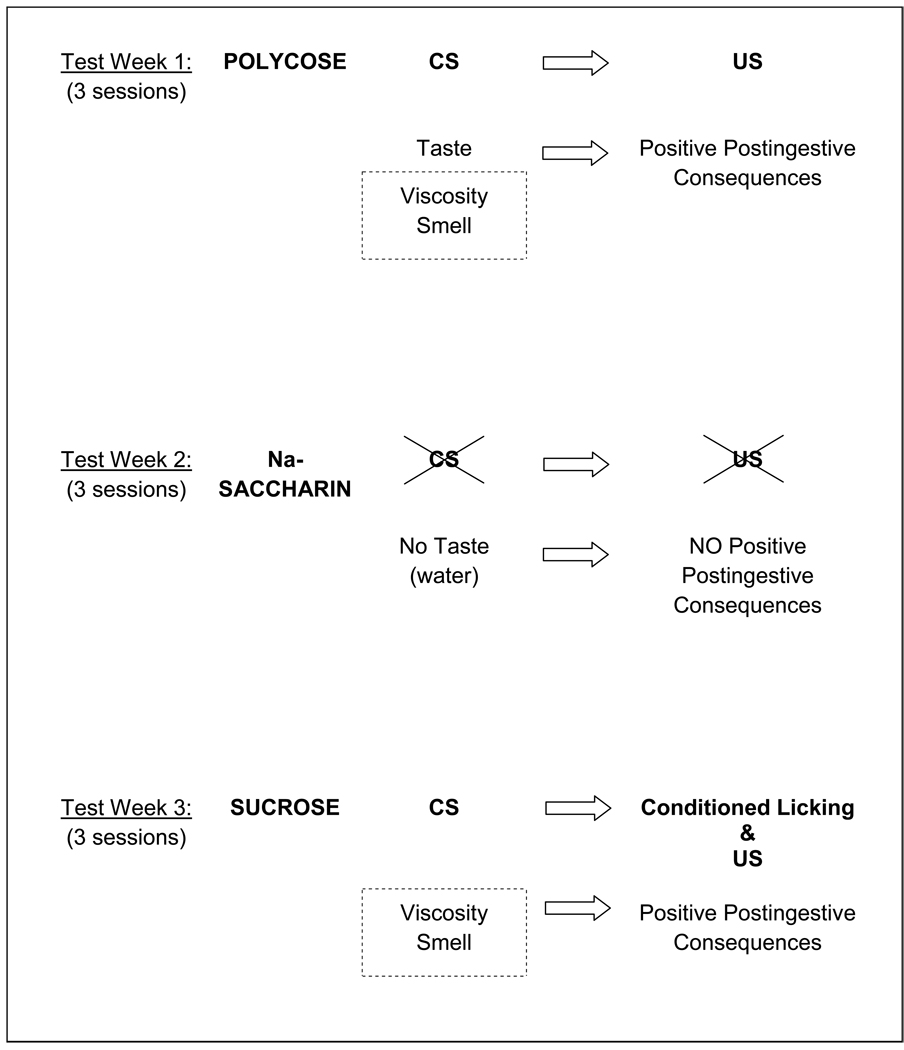
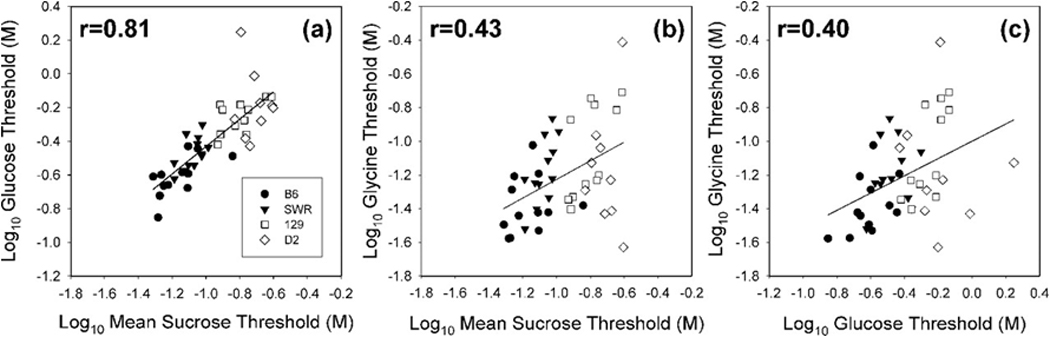
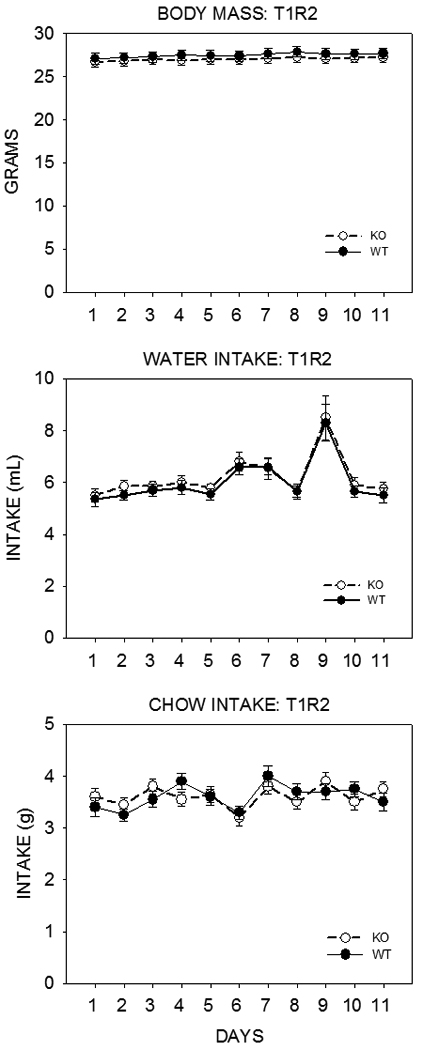
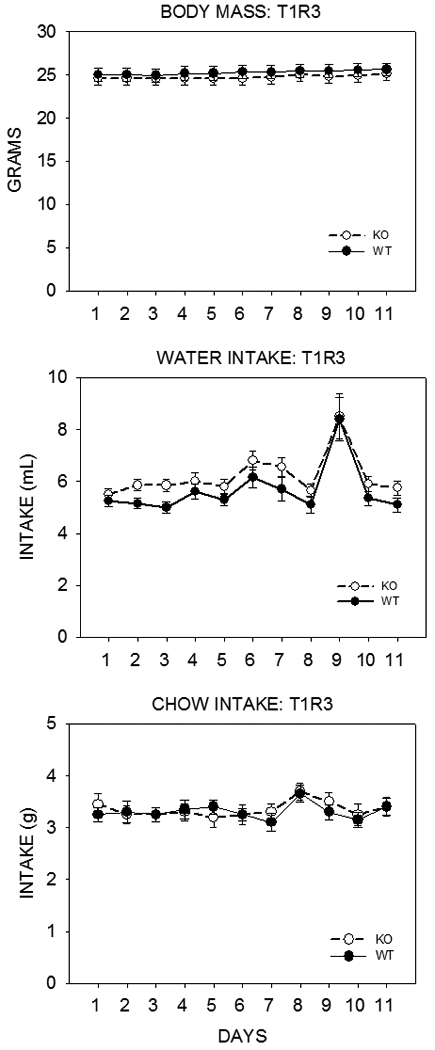
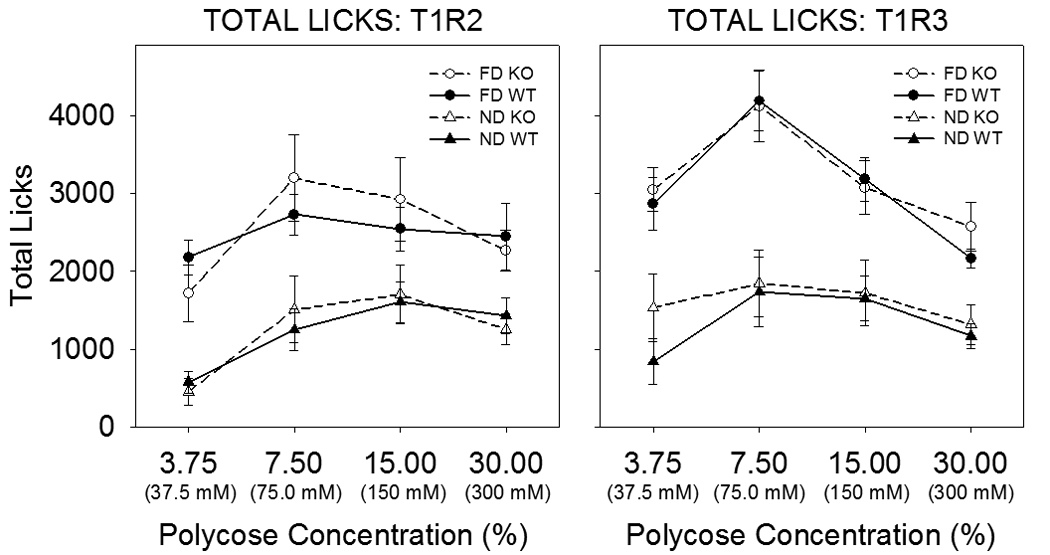
The discovery of the T1R family of Class C G protein-coupled receptors in the peripheral gustatory system a decade ago has been a tremendous advance for taste research, and its conceptual reach has extended to other organ systems. There are three proteins in the family, T1R1, T1R2, and T1R3, encoded by their respective genes, Tas1r1, Tas1r2, and Tas1r3. T1R2 combines with T1R3 to form a heterodimer that binds with sugars and other sweeteners. T1R3 also combines with T1R1 to form a heterodimer that binds with l-amino acids. These proteins are expressed not only in taste bud cells, but one or more of these T1Rs have also been identified in the nasal epithelium, gut, pancreas, liver, kidney, testes and brain in various mammalian species. Here we review current perspectives regarding the functional role of these receptors, concentrating on sweet taste and feeding. We also discuss behavioral findings suggesting that a glucose polymer mixture, Polycose, which rodents avidly prefer, appears to activate a receptor that does not depend on the combined expression of T1R2 and T1R3. In addition, although the T1Rs have been implicated as playing a role in glucose sensing, T1R2 knock-out (KO) and T1R3 KO mice display normal chow and fluid intake as well as normal body weight compared with same-sex littermate wild type (WT) controls. Moreover, regardless of whether they are fasted or not, these KO mice do not differ from their WT counterparts in their Polycose intake across a broad range of concentrations in 30-minute intake tests. The functional implications of these results and those in the literature are considered.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
T1R2 and T1R3 subunits are individually unnecessary for normal affective licking responses to Polycose: implications for saccharide taste receptors in mice.Am J Physiol Regul Integr Comp Physiol. 2009 Apr;296(4):R855-65. doi: 10.1152/ajpregu.90869.2008. Epub 2009 Jan 21. Am J Physiol Regul Integr Comp Physiol. 2009. PMID: 19158407 Free PMC article.
-
Electrophysiological responses to sugars and amino acids in the nucleus of the solitary tract of type 1 taste receptor double-knockout mice.J Neurophysiol. 2020 Feb 1;123(2):843-859. doi: 10.1152/jn.00584.2019. Epub 2020 Jan 8. J Neurophysiol. 2020. PMID: 31913749 Free PMC article.
-
Orosensory detection of sucrose, maltose, and glucose is severely impaired in mice lacking T1R2 or T1R3, but Polycose sensitivity remains relatively normal.Am J Physiol Regul Integr Comp Physiol. 2012 Jul 15;303(2):R218-35. doi: 10.1152/ajpregu.00089.2012. Epub 2012 May 23. Am J Physiol Regul Integr Comp Physiol. 2012. PMID: 22621968 Free PMC article.
-
Taste information derived from T1R-expressing taste cells in mice.Biochem J. 2016 Mar 1;473(5):525-36. doi: 10.1042/BJ20151015. Biochem J. 2016. PMID: 26912569 Review.
-
[Role of the sweet taste receptor in glucose metabolism: no sweets for diabetes?].Yakugaku Zasshi. 2015;135(6):763-7. doi: 10.1248/yakushi.14-00250-1. Yakugaku Zasshi. 2015. PMID: 26028411 Review. Japanese.
Cited by
-
A Novel Mechanism for T1R-Independent Taste Responses to Concentrated Sugars.J Neurosci. 2023 Feb 8;43(6):965-978. doi: 10.1523/JNEUROSCI.1760-22.2023. Epub 2023 Jan 9. J Neurosci. 2023. PMID: 36623875 Free PMC article.
-
Role of Taste Receptors as Sentinels of Innate Immunity in the Upper Airway.J Pathog. 2018 Oct 1;2018:9541987. doi: 10.1155/2018/9541987. eCollection 2018. J Pathog. 2018. PMID: 30363975 Free PMC article. Review.
-
Impact of obesity on taste receptor expression in extra-oral tissues: emphasis on hypothalamus and brainstem.Sci Rep. 2016 Jul 8;6:29094. doi: 10.1038/srep29094. Sci Rep. 2016. PMID: 27388805 Free PMC article.
-
Taste cell-expressed α-glucosidase enzymes contribute to gustatory responses to disaccharides.Proc Natl Acad Sci U S A. 2016 May 24;113(21):6035-40. doi: 10.1073/pnas.1520843113. Epub 2016 May 9. Proc Natl Acad Sci U S A. 2016. PMID: 27162343 Free PMC article.
-
Nutritive/non-nutritive sweeteners and high fat diet contribute to dysregulation of sweet taste receptors and metabolic derangements in oral, intestinal and central nervous tissues.Eur J Nutr. 2023 Dec;62(8):3149-3159. doi: 10.1007/s00394-023-03187-9. Epub 2023 Aug 3. Eur J Nutr. 2023. PMID: 37537344
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
