

Review
doi: 10.1104/pp.110.170399.
Epub 2011 Mar 4.
Regulation of starch biosynthesis in response to a fluctuating environment
Affiliations
- PMID: 21378102
- PMCID: PMC3091114
- DOI: 10.1104/pp.110.170399
Item in Clipboard
Review
Regulation of starch biosynthesis in response to a fluctuating environment
Plant Physiol.
2011 Apr.
No abstract available
Figures
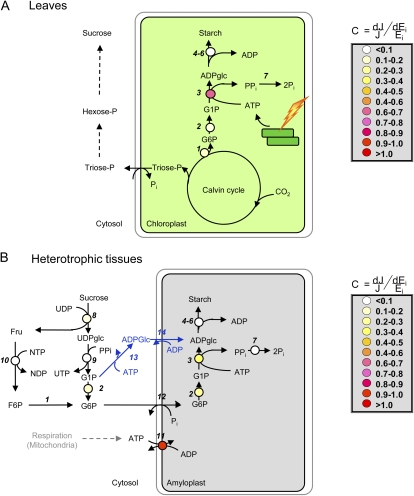
Schematic representation of the pathway of starch biosynthesis, its subcellular compartmentation, and distribution of flux control in photosynthetic leaves (A) and heterotrophic tissues (B). The reactions of the pathway of starch biosynthesis are catalyzed by the following enzymes: 1, phosphoglucoisomerase; 2, PGM; 3, AGPase; 4, SS; 5, SBE; 6, starch-debranching enzyme; 7, inorganic pyrophosphatase; 8, Suc synthase; 9 UDP-Glc pyrophosphorylase; 10, fructokinase; 11, ATP/ADP translocator; 12, Glc-6-P/Pi translocator; 13, cytosolic AGPase; and 14, ADP-Glc/ADP translocator (steps 13 and 14 are highlighted to be specific for cereal endosperm). The false color symbols represent the relative flux control coefficients (C) of the constituent enzymes, defined as the relation between the fractional change in enzyme activity (Ei) and starch flux (J). Missing symbols represent reactions for which experimental data are missing. Data were taken from Neuhaus and Stitt (1990) and Stitt et al. (2010) for leaves and from Geigenberger et al. (2004) for growing potato tubers.
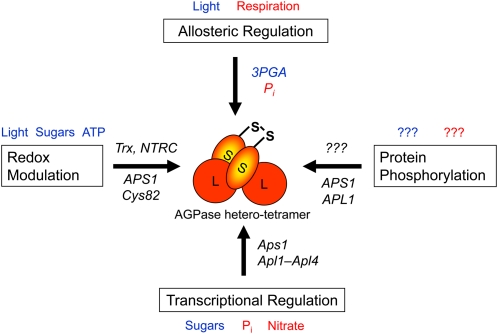
Regulation of plastidial AGPase by multiple mechanisms allows starch synthesis to respond across a range of time scales to a variety of physiological and environmental stimuli. Plastidial AGPase is a heterotetramer that contains two large (APL; 51 kD) and two slightly smaller (APS; 50 kD) subunits, which both have regulatory functions. Top, Allosteric regulation by 3PGA and Pi operates in a time frame of seconds to adjust the rate of starch synthesis to the balance between photosynthesis and Suc synthesis in leaves in the light and Suc breakdown and respiration in tubers. Left, Posttranslational redox modulation involves reversible disulfide bond formation between Cys-82 of the two small APS1 subunits, leading to changes in AGPase activity in response to light and sugar signals in a time frame of minutes to hours. The signaling components leading to redox modulation of AGPase involve Trx and NTRC, which are linked to photoreduced Fdx and interact with different sugar signals. Right, In Arabidopsis leaves, APS1 and APL1 have been identified as potential targets for reversible protein phosphorylation. More studies are needed to investigate the in vivo relevance of this mechanism and the underlying plastidial kinase network. Bottom, Transcriptional regulation in response to changes in carbon and nutrient supply allows more gradual changes in AGPase activity, which may require up to days to develop. Red font indicates inhibition, blue font indicates activation, and question marks indicate unknown (see main text for references).
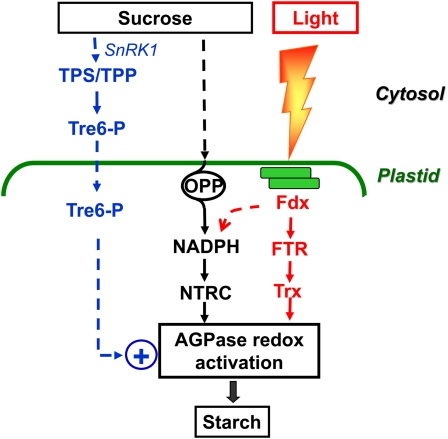
Posttranslational redox regulation of starch biosynthesis in response to light and sugar signals. Light activation of starch synthesis involves posttranslational redox activation of AGPase in the chloroplast (Hendriks et al., 2003) and resembles the light activation of enzymes of the Calvin-Benson cycle and related photosynthetic processes. Photosynthetic electron transport leads to reduction of Fdx, and reducing equivalents are transferred by FTR to Trx f or m, which activate target enzymes by the reduction of regulatory disulfides. NTRC, containing both an NADP-Trx reductase and a Trx in a single polypeptide, serves as an alternate system for transferring reducing equivalents from NADPH to AGPase, thereby enhancing storage starch synthesis (Michalska et al., 2009). In the light, NTRC is mainly linked to photoreduced Fdx via Fdx-NADP reductase (identified with the dashed arrow) and complements the FTR/Trx system in activating AGPase. In the dark or under conditions in which light reactions are impaired, NTRC is primarily linked to sugar oxidation via the initial reactions of the oxidative pentose phosphate pathway (OPP) and in this way regulates AGPase independently of the Fdx/Trx system. Redox activation of AGPase is also induced by Suc, which operates in leaves in the light and in nonphotosynthetic tissues (Tiessen et al., 2002; Hendriks et al., 2003). Tre-6-P acts an intracellular signal, linking Suc in the cytosol with AGPase in the plastid (Kolbe et al., 2005; Lunn et al., 2006). In the working model, an increase in Suc is sensed in the cytosol, leading to an increase in the level of Tre-6-P by modulating Tre-6-P synthase (TPS) and/or Tre-6-P phosphatase (TPP). Tre-6-P is taken up into the plastid and promotes NTRC- and/or FTR/Trx-dependent redox activation of AGPase by a yet unresolved mechanism. SnRK1 is also implicated in this Suc signaling pathway, although its specific role in signal transduction is not fully resolved yet (Tiessen et al., 2003; Jossier et al., 2009; Zhang et al., 2009). How SnRK1 and Tre-6-P interact in this signaling cascade is unclear and may depend on the tissue.
Similar articles
-
Redox regulation of carbon storage and partitioning in response to light and sugars.J Exp Bot. 2005 Jun;56(416):1469-79. doi: 10.1093/jxb/eri178. Epub 2005 Apr 29. J Exp Bot. 2005. PMID: 15863446 Review.
-
Sugar and ABA response pathways and the control of gene expression.Plant Cell Environ. 2006 Mar;29(3):426-34. doi: 10.1111/j.1365-3040.2005.01477.x. Plant Cell Environ. 2006. PMID: 17080596 Review.
-
Expression profiles of genes related to carbohydrate metabolism provide new insights into carbohydrate accumulation in seeds and seedlings of Ricinus communis in response to temperature.Plant Physiol Biochem. 2015 Oct;95:103-12. doi: 10.1016/j.plaphy.2015.07.023. Epub 2015 Jul 26. Plant Physiol Biochem. 2015. PMID: 26254183
-
Primary metabolism and nutrient assimilation: achieving a balanced diet.Curr Opin Plant Biol. 2003 Jun;6(3):205-7. doi: 10.1016/s1369-5266(03)00044-x. Curr Opin Plant Biol. 2003. PMID: 12753968 No abstract available.
-
Genome-wide analysis of starch metabolism genes in potato (Solanum tuberosum L.).BMC Genomics. 2017 Jan 5;18(1):37. doi: 10.1186/s12864-016-3381-z. BMC Genomics. 2017. PMID: 28056783 Free PMC article.
Cited by
-
Posttranslational Modification of Waxy to Genetically Improve Starch Quality in Rice Grain.Int J Mol Sci. 2021 May 3;22(9):4845. doi: 10.3390/ijms22094845. Int J Mol Sci. 2021. PMID: 34063649 Free PMC article. Review.
-
The Metabolic Reprogramming Induced by Sub-Optimal Nutritional and Light Inputs in Soilless Cultivated Green and Red Butterhead Lettuce.Int J Mol Sci. 2020 Sep 2;21(17):6381. doi: 10.3390/ijms21176381. Int J Mol Sci. 2020. PMID: 32887471 Free PMC article.
-
Leaf age and light stress affect the ability to diagnose P status in field grown potatoes.Front Plant Sci. 2023 Apr 20;14:1100318. doi: 10.3389/fpls.2023.1100318. eCollection 2023. Front Plant Sci. 2023. PMID: 37152149 Free PMC article.
-
Integrating roots into a whole plant network of flowering time genes in Arabidopsis thaliana.Sci Rep. 2016 Jun 29;6:29042. doi: 10.1038/srep29042. Sci Rep. 2016. PMID: 27352932 Free PMC article.
-
A fluorometric assay for trehalose in the picomole range.Plant Methods. 2013 Jun 20;9(1):21. doi: 10.1186/1746-4811-9-21. Plant Methods. 2013. PMID: 23786766 Free PMC article.
References
-
- ap Rees T, Hill SA. (1994) Metabolic control analysis of plant metabolism. Plant Cell Environ 17: 587–599
-
- Arsova B, Hoja U, Wimmelbacher M, Greiner E, Ustün S, Melzer M, Petersen K, Lein W, Börnke F. (2010) Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 22: 1498–1515 - PMC - PubMed
-
- Baena-González E, Rolland F, Thevelein JM, Sheen J. (2007) A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942 - PubMed
-
- Baginsky S, Gruissem W. (2009) The chloroplast kinase network: new insights from large-scale phosphoproteome profiling. Mol Plant 2: 1141–1153 - PubMed
-
- Ball SG, Morell MK. (2003) From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule. Annu Rev Plant Biol 54: 207–233 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
