

Massive production of small RNAs from a non-coding region of Cauliflower mosaic virus in plant defense and viral counter-defense
- PMID: 21378120
- PMCID: PMC3130284
- DOI: 10.1093/nar/gkr119
Massive production of small RNAs from a non-coding region of Cauliflower mosaic virus in plant defense and viral counter-defense
Abstract
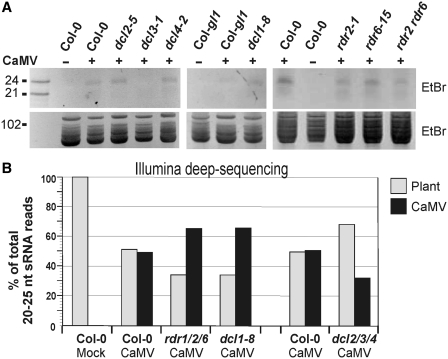
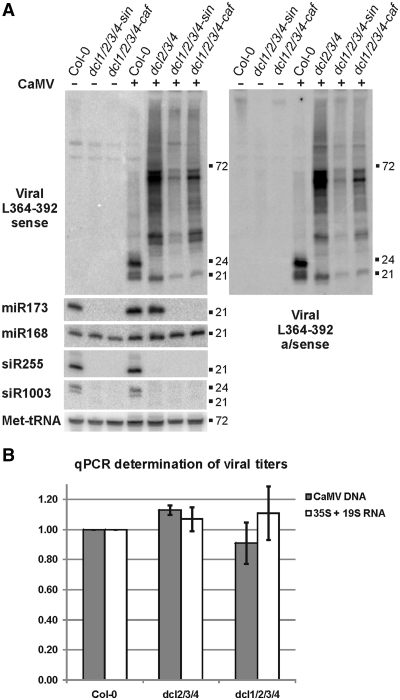
To successfully infect plants, viruses must counteract small RNA-based host defense responses. During infection of Arabidopsis, Cauliflower mosaic pararetrovirus (CaMV) is transcribed into pregenomic 35S and subgenomic 19S RNAs. The 35S RNA is both reverse transcribed and also used as an mRNA with highly structured 600 nt leader. We found that this leader region is transcribed into long sense- and antisense-RNAs and spawns a massive quantity of 21, 22 and 24 nt viral small RNAs (vsRNAs), comparable to the entire complement of host-encoded small-interfering RNAs and microRNAs. Leader-derived vsRNAs were detected bound to the Argonaute 1 (AGO1) effector protein, unlike vsRNAs from other viral regions. Only negligible amounts of leader-derived vsRNAs were bound to AGO4. Genetic evidence showed that all four Dicer-like (DCL) proteins mediate vsRNA biogenesis, whereas the RNA polymerases Pol IV, Pol V, RDR1, RDR2 and RDR6 are not required for this process. Surprisingly, CaMV titers were not increased in dcl1/2/3/4 quadruple mutants that accumulate only residual amounts of vsRNAs. Ectopic expression of CaMV leader vsRNAs from an attenuated geminivirus led to increased accumulation of this chimeric virus. Thus, massive production of leader-derived vsRNAs does not restrict viral replication but may serve as a decoy diverting the silencing machinery from viral promoter and coding regions.
Figures




References
-
- Chapman EJ, Carrington JC. Specialization and evolution of endogenous small RNA pathways. Nat. Rev. Genet. 2007;8:884–896. - PubMed
-
- Ruiz-Ferrer V, Voinnet O. Roles of plant small RNAs in biotic stress responses. Annu. Rev. Plant Biol. 2009;60:485–510. - PubMed
-
- Hamilton AJ, Baulcombe DC. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science. 1999;286:950–952. - PubMed
-
- Csorba T, Pantaleo V, Burgyán J. RNA silencing: an antiviral mechanism. Adv. Virus Res. 2009;75:35–71. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
