

7,3',4'-Trihydroxyisoflavone, a metabolite of the soy isoflavone daidzein, suppresses ultraviolet B-induced skin cancer by targeting Cot and MKK4
- PMID: 21378167
- PMCID: PMC3077626
- DOI: 10.1074/jbc.M110.147348
7,3',4'-Trihydroxyisoflavone, a metabolite of the soy isoflavone daidzein, suppresses ultraviolet B-induced skin cancer by targeting Cot and MKK4
Abstract
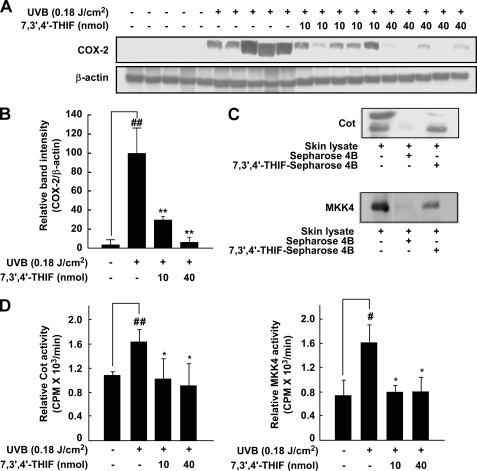
Nonmelanoma skin cancer is one of the most frequently occurring cancers in the United States. Chronic exposure to UVB irradiation is a major cause of this cancer. Daidzein, along with genistein, is a major isoflavone found in soybeans; however, little is known about the chemopreventive effects of daidzein and its metabolites in UVB-induced skin cancer. Here, we found that 7,3',4'-trihydroxyisoflavone (THIF), a major metabolite of daidzein, effectively inhibits UVB-induced cyclooxygenase 2 (COX-2) expression through the inhibition of NF-κB transcription activity in mouse skin epidermal JB6 P+ cells. In contrast, daidzein had no effect on COX-2 expression levels. Data from Western blot and kinase assays showed that 7,3',4'-THIF inhibited Cot and MKK4 activity, thereby suppressing UVB-induced phosphorylation of mitogen-activated protein kinases. Pull-down assays indicated that 7,3',4'-THIF competed with ATP to inhibit Cot or MKK4 activity. Topical application of 7,3',4'-THIF clearly suppressed the incidence and multiplicity of UVB-induced tumors in hairless mouse skin. Hairless mouse skin results also showed that 7,3',4'-THIF inhibits Cot or MKK4 kinase activity directly, resulting in suppressed UVB-induced COX-2 expression. A docking study revealed that 7,3',4'-THIF, but not daidzein, easily docked to the ATP binding site of Cot and MKK4, which is located between the N- and C-lobes of the kinase domain. Collectively, these results provide insight into the biological actions of 7,3',4'-THIF, a potential skin cancer chemopreventive agent.
Figures





References
-
- Athar M., An K. P., Morel K. D., Kim A. L., Aszterbaum M., Longley J., Epstein E. H., Jr., Bickers D. R. (2001) Biochem. Biophys. Res. Commun. 280, 1042–1047 - PubMed
-
- Rundhaug J. E., Fischer S. M. (2008) Photochem. Photobiol. 84, 322–329 - PubMed
-
- Katiyar S. K., Korman N. J., Mukhtar H., Agarwal R. (1997) J. Natl. Cancer Inst. 89, 556–566 - PubMed
-
- Buckman S. Y., Gresham A., Hale P., Hruza G., Anast J., Masferrer J., Pentland A. P. (1998) Carcinogenesis 19, 723–729 - PubMed
-
- Marwaha V., Chen Y. H., Helms E., Arad S., Inoue H., Bord E., Kishore R., Sarkissian R. D., Gilchrest B. A., Goukassian D. A. (2005) J. Biol. Chem. 280, 32379–32388 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
