

Sex-specific activation of cell death signalling pathways in cerebellar granule neurons exposed to oxygen glucose deprivation followed by reoxygenation
- PMID: 21382016
- PMCID: PMC3072765
- DOI: 10.1042/AN20100032
Sex-specific activation of cell death signalling pathways in cerebellar granule neurons exposed to oxygen glucose deprivation followed by reoxygenation
Abstract
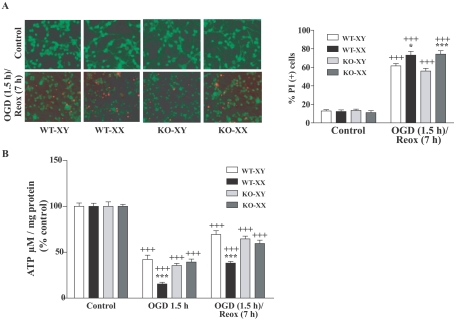
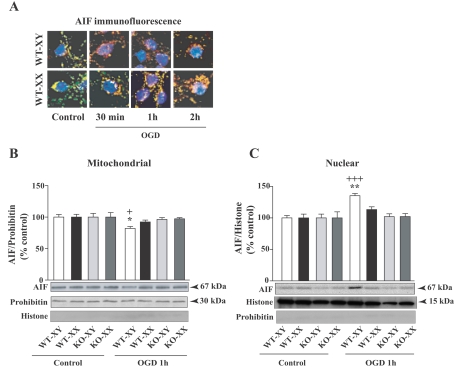
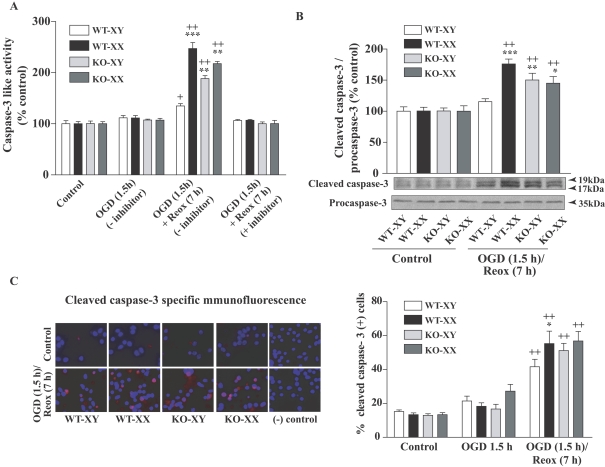
Neuronal death pathways following hypoxia-ischaemia are sexually dimorphic, but the underlying mechanisms are unclear. We examined cell death mechanisms during OGD (oxygen-glucose deprivation) followed by Reox (reoxygenation) in segregated male (XY) and female (XX) mouse primary CGNs (cerebellar granule neurons) that are WT (wild-type) or Parp-1 [poly(ADP-ribose) polymerase 1] KO (knockout). Exposure of CGNs to OGD (1.5 h)/Reox (7 h) caused cell death in XY and XX neurons, but cell death during Reox was greater in XX neurons. ATP levels were significantly lower after OGD/Reox in WT-XX neurons than in XY neurons; this difference was eliminated in Parp-1 KO-XX neurons. AIF (apoptosis-inducing factor) was released from mitochondria and translocated to the nucleus by 1 h exclusively in WT-XY neurons. In contrast, there was a release of Cyt C (cytochrome C) from mitochondria in WT-XX and Parp-1 KO neurons of both sexes; delayed activation of caspase 3 was observed in the same three groups. Thus deletion of Parp-1 shunted cell death towards caspase 3-dependent apoptosis. Delayed activation of caspase 8 was also observed in all groups after OGD/Reox, but was much greater in XX neurons, and caspase 8 translocated to the nucleus in XX neurons only. Caspase 8 activation may contribute to increased XX neuronal death during Reox, via caspase 3 activation. Thus, OGD/Reox induces death of XY neurons via a PARP-1-AIF-dependent mechanism, but blockade of PARP-1-AIF pathway shifts neuronal death towards a caspase-dependent mechanism. In XX neurons, OGD/Reox caused prolonged depletion of ATP and delayed activation of caspase 8 and caspase 3, culminating in greater cell death during Reox.
Figures



Similar articles
-
Sex differences in mitochondrial biogenesis determine neuronal death and survival in response to oxygen glucose deprivation and reoxygenation.BMC Neurosci. 2014 Jan 10;15:9. doi: 10.1186/1471-2202-15-9. BMC Neurosci. 2014. PMID: 24410996 Free PMC article.
-
Mitochondrial impairment induced by poly(ADP-ribose) polymerase-1 activation in cortical neurons after oxygen and glucose deprivation.J Neurochem. 2005 Oct;95(1):179-90. doi: 10.1111/j.1471-4159.2005.03353.x. J Neurochem. 2005. PMID: 16181422
-
Poly(ADP-ribose) polymerase (PARP)-1-independent apoptosis-inducing factor (AIF) release and cell death are induced by eleostearic acid and blocked by alpha-tocopherol and MEK inhibition.J Biol Chem. 2010 Apr 23;285(17):13079-91. doi: 10.1074/jbc.M109.044206. Epub 2010 Feb 22. J Biol Chem. 2010. PMID: 20177052 Free PMC article.
-
Deadly conversations: nuclear-mitochondrial cross-talk.J Bioenerg Biomembr. 2004 Aug;36(4):287-94. doi: 10.1023/B:JOBB.0000041755.22613.8d. J Bioenerg Biomembr. 2004. PMID: 15377859 Review.
-
Poly(ADP-ribose) signals to mitochondrial AIF: a key event in parthanatos.Exp Neurol. 2009 Aug;218(2):193-202. doi: 10.1016/j.expneurol.2009.03.020. Epub 2009 Mar 28. Exp Neurol. 2009. PMID: 19332058 Free PMC article. Review.
Cited by
-
ADP-ribosylation: from molecular mechanisms to human disease.Genet Mol Biol. 2019 Dec 13;43(1 suppl 1):e20190075. doi: 10.1590/1678-4685-GMB-2019-0075. eCollection 2019. Genet Mol Biol. 2019. PMID: 31930280 Free PMC article.
-
Cell death atlas of the postnatal mouse ventral forebrain and hypothalamus: effects of age and sex.J Comp Neurol. 2013 Aug 1;521(11):2551-69. doi: 10.1002/cne.23298. J Comp Neurol. 2013. PMID: 23296992 Free PMC article.
-
The aldehyde group of gossypol induces mitochondrial apoptosis via ROS-SIRT1-p53-PUMA pathway in male germline stem cell.Oncotarget. 2017 Oct 24;8(59):100128-100140. doi: 10.18632/oncotarget.22044. eCollection 2017 Nov 21. Oncotarget. 2017. PMID: 29245965 Free PMC article.
-
Mesenchymal stem cells protect neurons against hypoxic-ischemic injury via inhibiting parthanatos, necroptosis, and apoptosis, but not autophagy.Cell Mol Neurobiol. 2017 Mar;37(2):303-313. doi: 10.1007/s10571-016-0370-3. Epub 2016 Apr 4. Cell Mol Neurobiol. 2017. PMID: 27044018 Free PMC article.
-
Mitochondria and sensory processing in inflammatory and neuropathic pain.Front Pain Res (Lausanne). 2022 Oct 17;3:1013577. doi: 10.3389/fpain.2022.1013577. eCollection 2022. Front Pain Res (Lausanne). 2022. PMID: 36324872 Free PMC article. Review.
References
-
- Ame JC, Rolli V, Schreiber V, Niedergang C, Apiou F, Decker P, Muller S, Hoger T, Menissier-de Murcia J, de Murcia G. PARP-2, A novel mammalian DNA damage-dependent poly (ADP-ribose) polymerase. J Biol Chem. 1999;274:17860–17868. - PubMed
-
- Arnold AP, Xu J, Grisham W, Chen X, Kim YH, Itoh Y. Minireview: sex chromosomes and brain sexual differentiation. Endocrinology. 2004;145:1057–1062. - PubMed
-
- Barks JD, Silverstein FS. Excitatory amino acids contribute to the pathogenesis of perinatal hypoxic-ischemic brain injury. Brain Pathol. 1992;2:235–243. - PubMed
-
- Benchoua A, Couriaud C, Guegan C, Tartier L, Couvert P, Friocourt G, Chelly J, Menissier-de Murcia J, Onteniente B. Active caspase-8 translocates into the nucleus of apoptotic cells to inactivate poly (ADP-ribose) polymerase-2. J Biol Chem. 2002;277:34217–34222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous