

Development of synaptic transmission to respiratory motoneurons
- PMID: 21382524
- PMCID: PMC3157583
- DOI: 10.1016/j.resp.2011.03.002
Development of synaptic transmission to respiratory motoneurons
Abstract
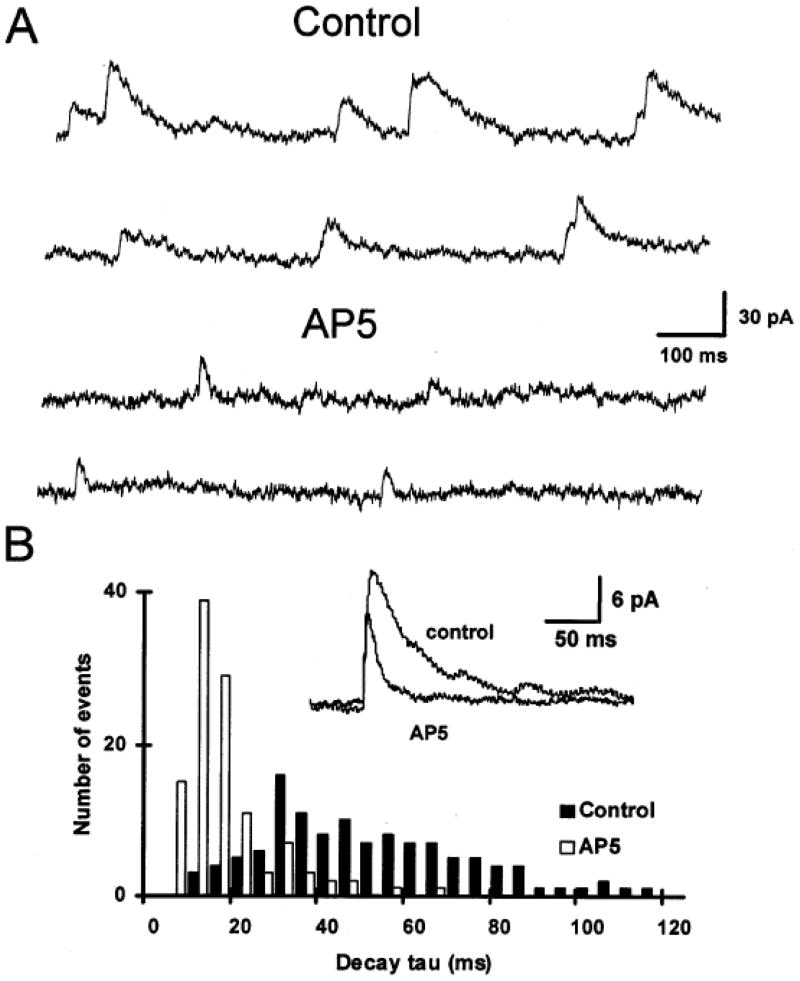
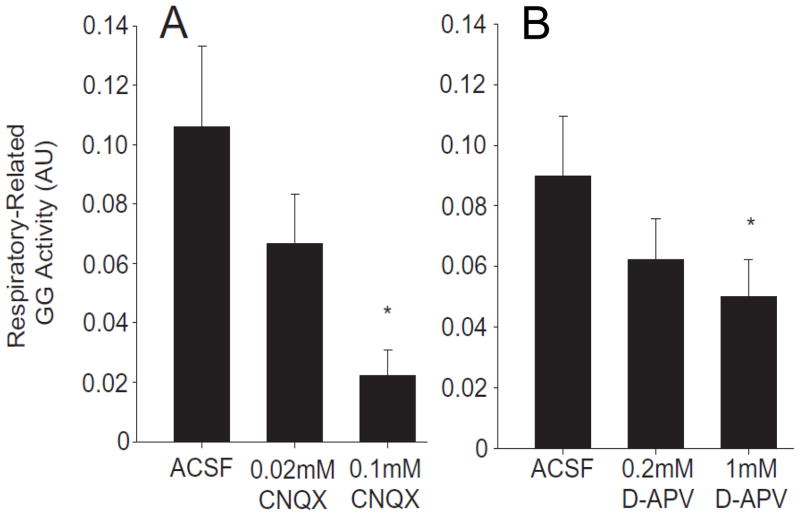
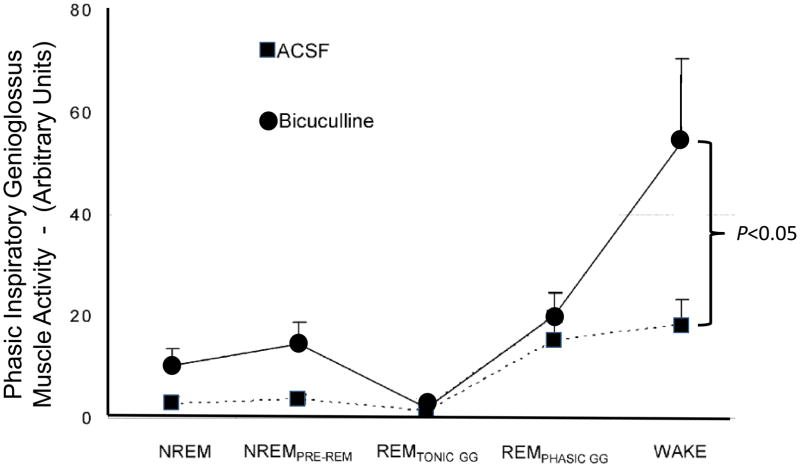
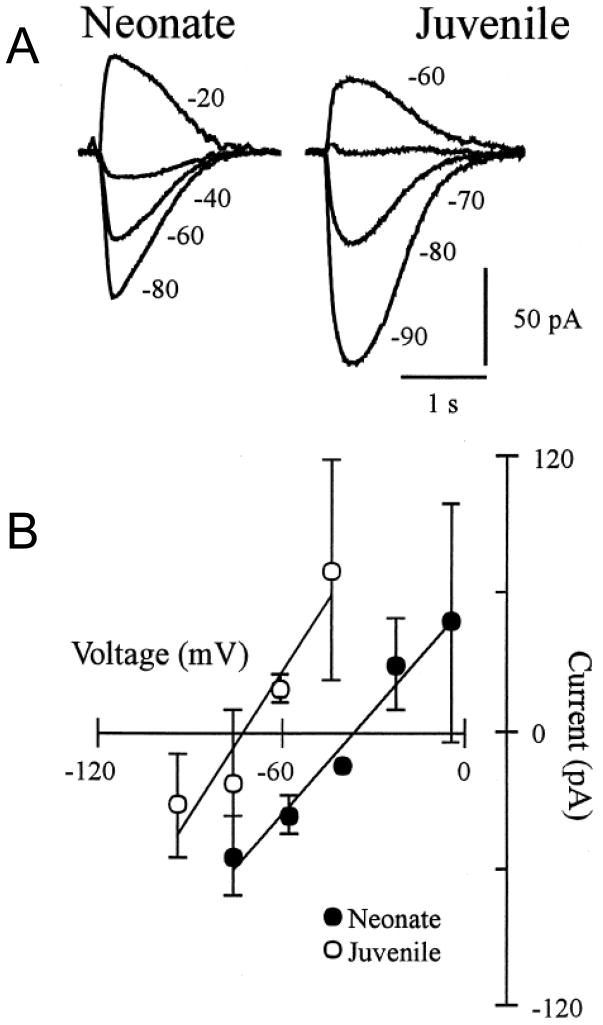
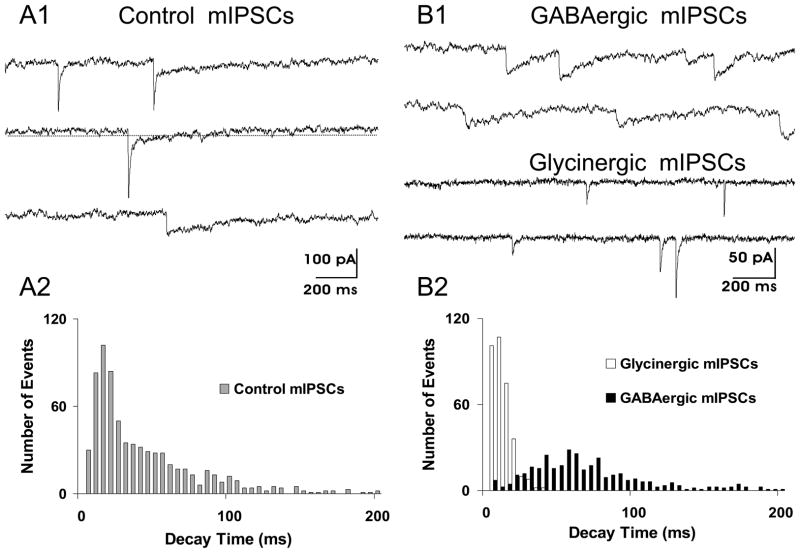
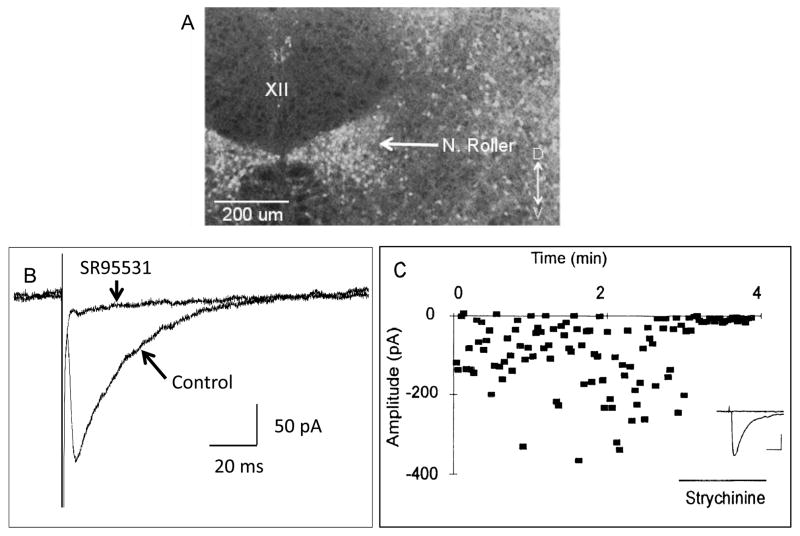
Respiratory motoneurons provide the exclusive drive to respiratory muscles and therefore are a key relay between brainstem neural circuits that generate respiratory rhythm and respiratory muscles that control moment of gases into and out of the airways and lungs. This review is focused on postnatal development of fast ionotropic synaptic transmission to respiratory motoneurons, with a focus on hypoglossal motoneurons (HMs). Glutamatergic synaptic transmission to HMs involves activation of both non-NMDA and NMDA receptors and during the postnatal period co-activation of these receptors located at the same synapse may occur. Further, the relative role of each receptor type in inspiratory-phase motoneuron depolarization is dependent on the type of preparation used (in vitro versus in vivo; neonatal versus adult). Respiratory motoneurons receive both glycinergic and GABAergic inhibitory synaptic inputs. During inspiration phrenic and HMs receive concurrent excitatory and inhibitory synaptic inputs. During postnatal development in HMs GABAergic and glycinergic synaptic inputs have slow kinetics and are depolarizing and with postnatal development they become faster and hyperpolarizing. Additionally shunting inhibition may play an important role in synaptic processing by respiratory motoneurons.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
References
-
- Aldes LD, Chronister RB, Marco LA. Distribution of glutamic acid decarboxylase and gamma-aminobutyric acid in the hypoglossal nucleus in the rat. J Neurosci Res. 1988;19:343–348. - PubMed
-
- Altmann H, ten Bruggencate G, Sonnhof U. Differential strength of action of glycine and GABA in hypoglossus nucleus. Pflugers Arch. 1972;331:90–94. - PubMed
-
- Altschuler SM, Bao X, Miselis RR. Dendritic architecture of hypoglossal motoneurons projecting to extrinsic tongue musculature in the rat. J Comp Neurol. 1994;342:538–550. - PubMed
-
- Baker SN, Kliner JN, Pinches EM, Lemon RN. The role of synchrony and oscillations in the motor output. Exp Brain Res. 1999;128:109–117. - PubMed
-
- Ben-Ari Y, Gaiarsa JL, Tyzio R, Khazipov R. GABA: A pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev. 2007;87:1215–1284. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
