

Phosphorylation regulates OLIG2 cofactor choice and the motor neuron-oligodendrocyte fate switch
- PMID: 21382552
- PMCID: PMC3093612
- DOI: 10.1016/j.neuron.2011.01.030
Phosphorylation regulates OLIG2 cofactor choice and the motor neuron-oligodendrocyte fate switch
Abstract
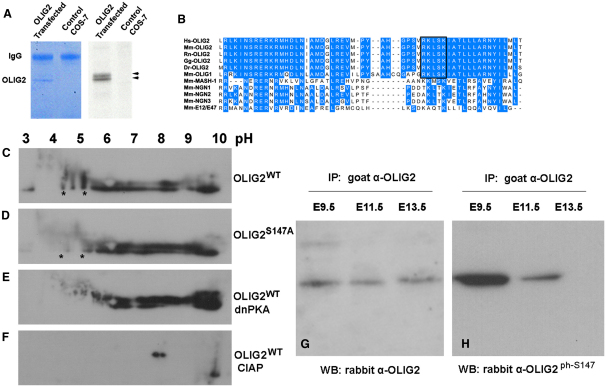
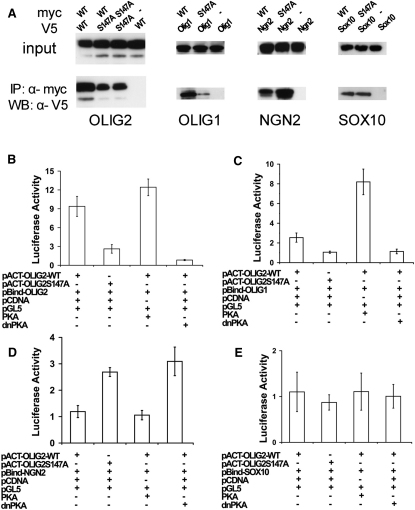
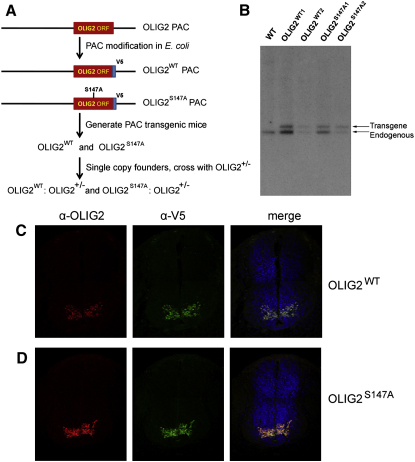
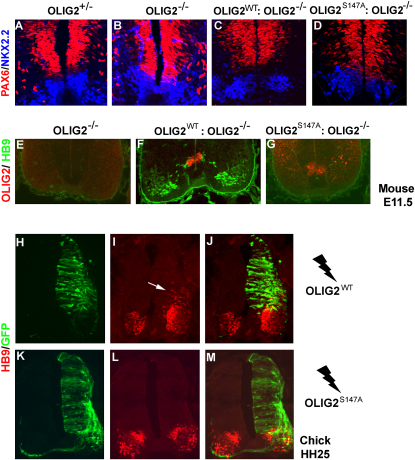
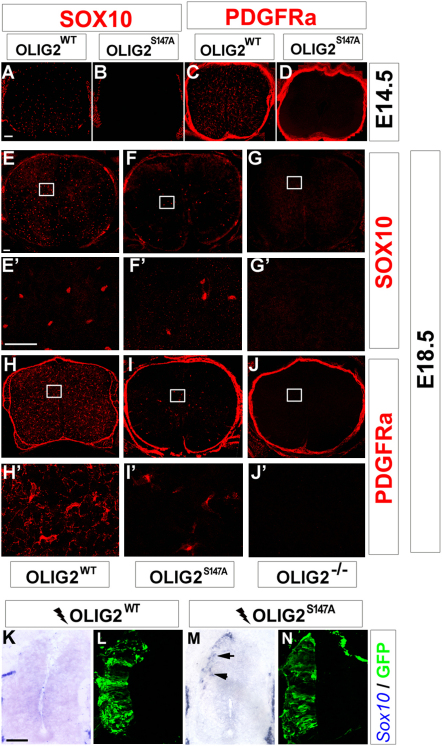
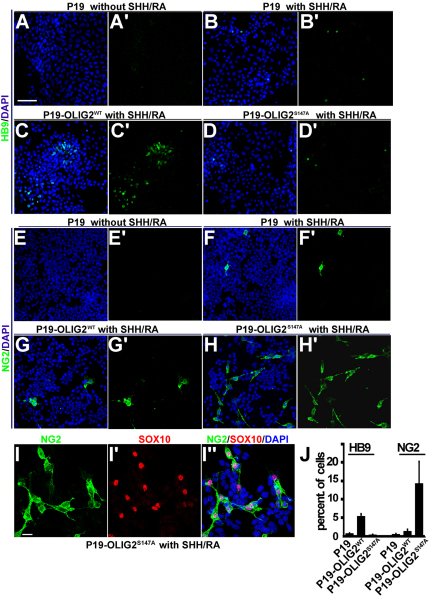
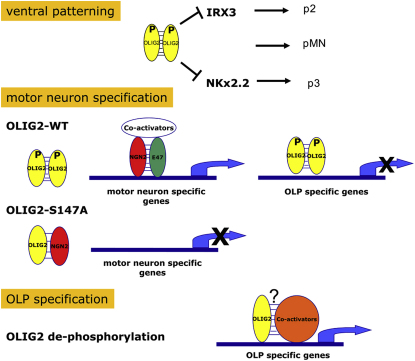
A fundamental feature of central nervous system development is that neurons are generated before glia. In the embryonic spinal cord, for example, a group of neuroepithelial stem cells (NSCs) generates motor neurons (MNs), before switching abruptly to oligodendrocyte precursors (OLPs). We asked how transcription factor OLIG2 participates in this MN-OLP fate switch. We found that Serine 147 in the helix-loop-helix domain of OLIG2 was phosphorylated during MN production and dephosphorylated at the onset of OLP genesis. Mutating Serine 147 to Alanine (S147A) abolished MN production without preventing OLP production in transgenic mice, chicks, or cultured P19 cells. We conclude that S147 phosphorylation, possibly by protein kinase A, is required for MN but not OLP genesis and propose that dephosphorylation triggers the MN-OLP switch. Wild-type OLIG2 forms stable homodimers, whereas mutant (unphosphorylated) OLIG2(S147A) prefers to form heterodimers with Neurogenin 2 or other bHLH partners, suggesting a molecular basis for the switch.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
All the embryo's a stage, and Olig2 in its time plays many parts.Neuron. 2011 Mar 10;69(5):833-5. doi: 10.1016/j.neuron.2011.02.037. Neuron. 2011. PMID: 21382543
-
Development: directing development through phosphorylation.Nat Rev Neurosci. 2011 May;12(5):248-9. doi: 10.1038/nrn3030. Epub 2011 Apr 13. Nat Rev Neurosci. 2011. PMID: 21505513 No abstract available.
References
-
- Arnett H.A., Fancy S.P., Alberta J.A., Zhao C., Plant S.R., Kaing S., Raine C.S., Rowitch D.H., Franklin R.J., Stiles C.D. bHLH transcription factor Olig1 is required to repair demyelinated lesions in the CNS. Science. 2004;306:2111–2115. - PubMed
-
- Bai C.B., Stephen D., Joyner A.L. All mouse ventral spinal cord patterning by hedgehog is Gli dependent and involves an activator function of Gli3. Dev. Cell. 2004;6:103–115. - PubMed
-
- Berg R.W., McBurney M.W. Cell density and cell cycle effects on retinoic acid-induced embryonal carcinoma cell differentiation. Dev. Biol. 1990;138:123–135. - PubMed
-
- Blom N., Gammeltoft S., Brunak S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999;294:1351–1362. - PubMed
-
- Blom N., Sicheritz-Ponten T., Gupta R., Gammeltoft S., Brunak S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics. 2004;4:1633–1649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
