

Effect of cytokine interplay on macrophage polarization during chronic pulmonary infection with Cryptococcus neoformans
- PMID: 21383052
- PMCID: PMC3088136
- DOI: 10.1128/IAI.01270-10
Effect of cytokine interplay on macrophage polarization during chronic pulmonary infection with Cryptococcus neoformans
Abstract
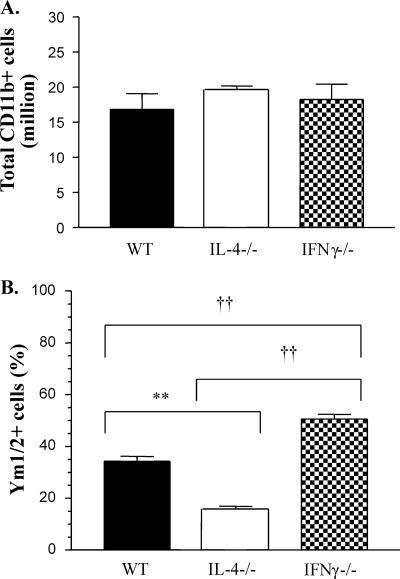
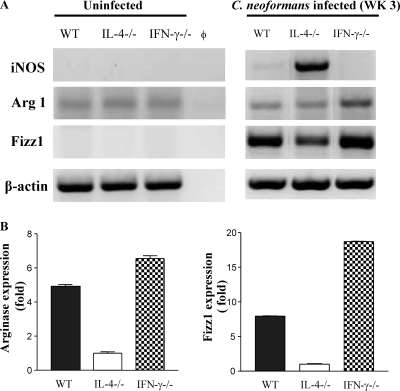
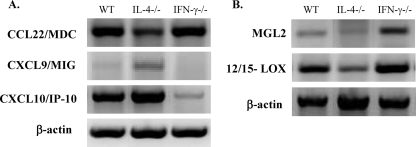
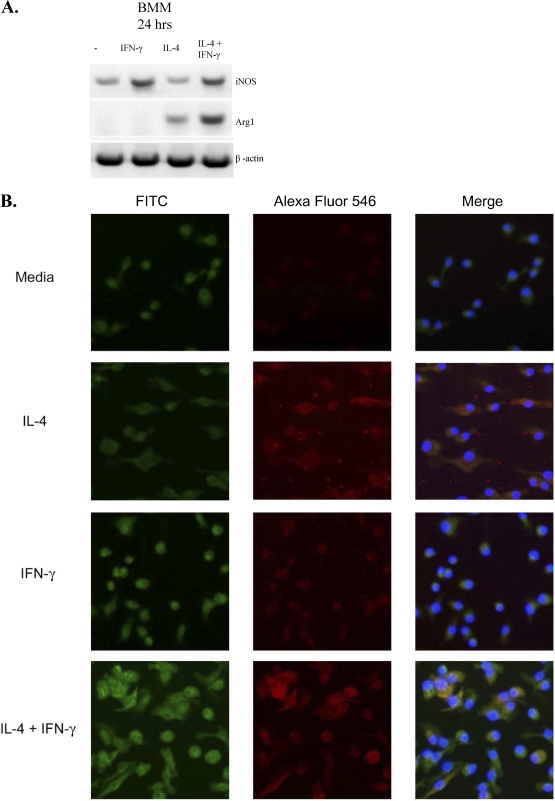
The immune response to Cryptococcus neoformans following pulmonary infection of C57BL/6 wild-type (WT) mice results in the development of persistent infection with characteristics of allergic bronchopulmonary mycosis (ABPM). To further clarify the role of Th1/Th2 polarizing cytokines in this model, we performed kinetic analysis of cytokine responses and compared cytokine profiles, pathologies, and macrophage (Mac) polarization status in C. neoformans-infected WT, interleukin-4-deficient (IL-4(-/-)), and gamma interferon-deficient (IFN-γ(-/-)) C57BL/6 mice. Results show that cytokine expression in the infected WT mice is not permanently Th2 biased but changes dynamically over time. Using multiple Mac activation markers, we further demonstrate that IL-4 and IFN-γ regulate the polarization state of Macs in this model. A higher IL-4/IFN-γ ratio leads to the development of alternatively activated Macs (aaMacs), whereas a higher IFN-γ/IL-4 ratio leads to the generation of classically activated Macs (caMacs). WT mice that coexpress IL-4 and IFN-γ during fungal infection concurrently display both types of Mac polarization markers. Concurrent stimulation of Macs with IFN-γ and IL-4 results in an upregulation of both sets of markers within the same cells, i.e., formation of an intermediate aaMac/caMac phenotype. These cells express both inducible nitric oxide synthase (important for clearance) and arginase (associated with chronic/progressive infection). Together, our data demonstrate that the interplay between Th1 and Th2 cytokines supports chronic infection, chronic inflammation, and the development of ABPM pathology in C. neoformans-infected lungs. This cytokine interplay modulates Mac differentiation, including generation of an intermediate caMac/aaMac phenotype, which in turn may support chronic "steady-state" fungal infection and the resultant ABPM pathology.
Figures
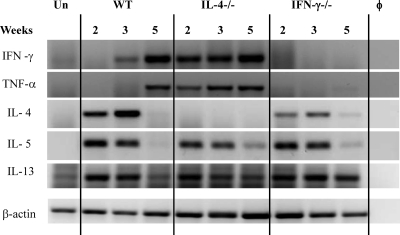
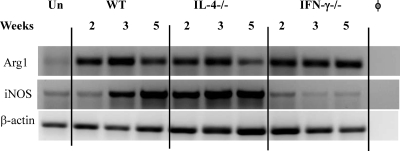
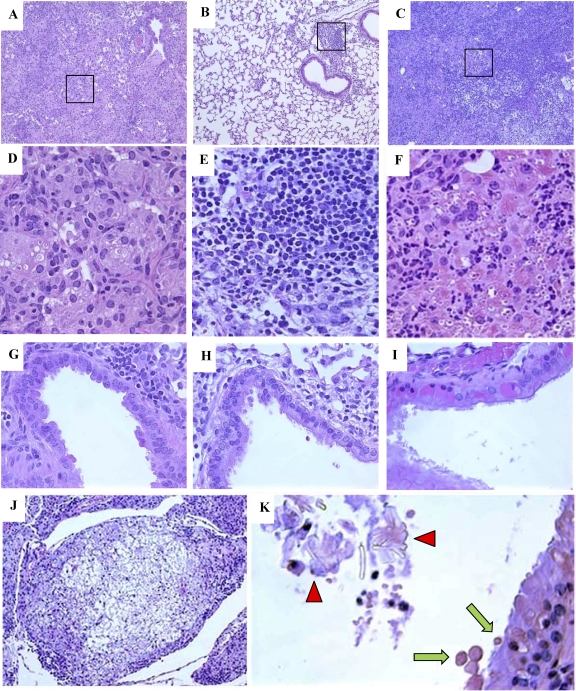
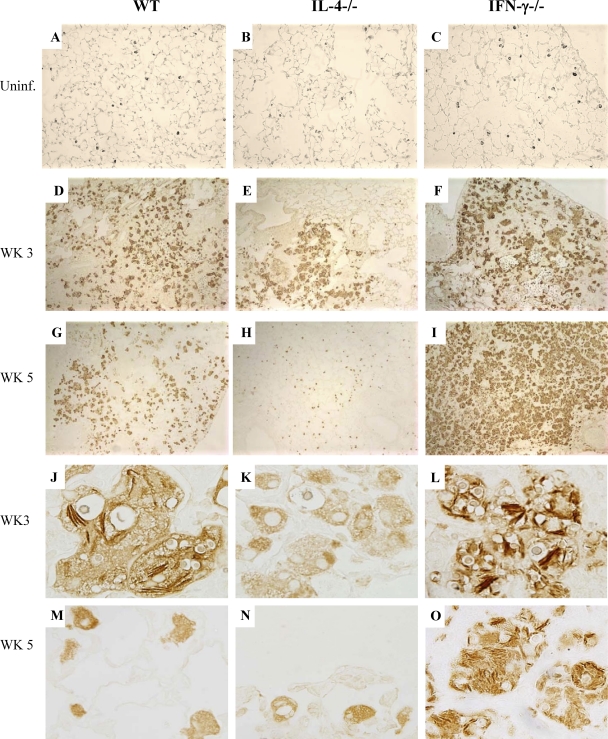
References
-
- Arora S., et al. 2005. Role of IFN-gamma in regulating T2 immunity and the development of alternatively activated macrophages during allergic bronchopulmonary mycosis. J. Immunol. 174:6346–6356 - PubMed
-
- Brys L., et al. 2005. Reactive oxygen species and 12/15-lipoxygenase contribute to the antiproliferative capacity of alternatively activated myeloid cells elicited during helminth infection. J. Immunol. 174:6095–6104 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
