

Review
doi: 10.1083/jcb.201010129.
On emerging nuclear order
Affiliations
- PMID: 21383074
- PMCID: PMC3051810
- DOI: 10.1083/jcb.201010129
Item in Clipboard
Review
On emerging nuclear order
J Cell Biol.
.
Abstract
Although the nonrandom nature of interphase chromosome arrangement is widely accepted, how nuclear organization relates to genomic function remains unclear. Nuclear subcompartments may play a role by offering rich microenvironments that regulate chromatin state and ensure optimal transcriptional efficiency. Technological advances now provide genome-wide and four-dimensional analyses, permitting global characterizations of nuclear order. These approaches will help uncover how seemingly separate nuclear processes may be coupled and aid in the effort to understand the role of nuclear organization in development and disease.
Figures
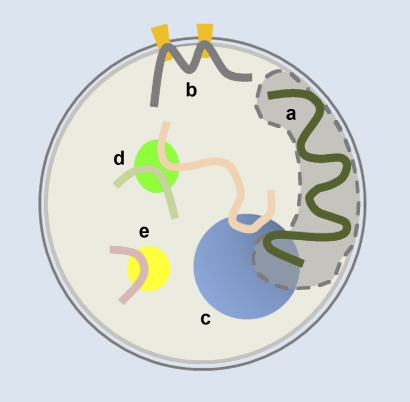
Chromosome conformation and transcriptional activity are affected by the association of chromosomal regions with peripheral or central subcompartments. (a) The nuclear lamina and adjacent nuclear space, (b) nuclear pore complexes, (c) a nucleolus, where ribosomal DNA loci from different chromosomes cluster, (d) a transcription factory, where coregulated genes preferentially colocalize, and (e) a polycomb body. The filled gray region represents a CT that is interacting with both a and c.
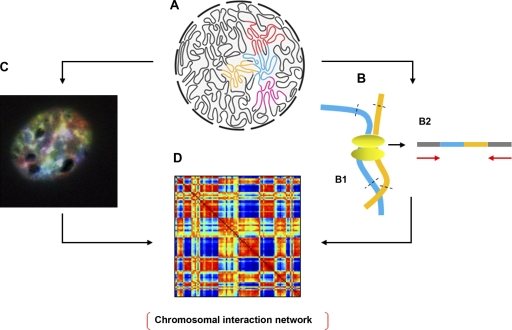
Methods for exploring the 3D genome. (A) A schematic representation of the 3D genome (courtesy of J. Dekker and N.L. van Berkum, University of Massachusetts Medical School, Worcester, MA). (B) The Hi-C technique. (B1) DNA is cross-linked and digested with restriction enzymes. (B2) High throughput sequencing (8.5 million reads) is used to determine the spatial proximity of sequences, including those on the same or different chromosomes using paired-end sequencing. (C) Image of a murine hematopoietic progenitor interphase nucleus labeled by SKY. All chromosomes are labeled with a unique color to visualize their territories. (D) Once algorithms are developed to characterize the spatial relationships among all CTs in the interphase nucleus simultaneously from SKY data, spatial proximity maps can be generated that integrate both SKY and Hi-C data to more precisely define inter- and intrachromosomal interactions.
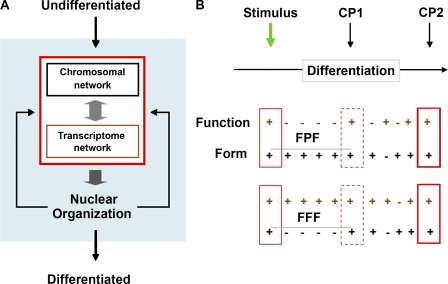
The mechanics of self-organization and differentiation. (A) Local interactions that make up the transcriptome network (gene coregulation) and spatial architecture that makes up the chromosomal network mutually influence each other and lead to the emergence of cell-specific nuclear organization. In turn, this order feeds back to strengthen the local associations, and the self-organized system fine tunes over time. (B) Two possible models, form precedes function (FPF) and form follows function (FFF), representing the relationship between chromosomal (form) and transcriptome (function) networks over time during differentiation (horizontal black arrow). At the green arrow, a stimulus initiates the process of differentiation. CP1 and CP2 are critical points 1 and 2 or defined states in nuclear architecture. The solid, dotted, and bold boxes represent nuclear organization as determined by the interaction between the networks. Plus and minus symbols show changing or unchanging architecture, respectively, within each network. In the undifferentiated state in both models, the transcription and space networks are changing and mutually influence each other. After the stimulus in the form precedes function model, only the space network changes over time until CP1. At CP1, when the space network has reached a particular configuration, the transcription network begins to change, and again they become mutually related. In the form follows function model, only the transcription network changes over time until CP1, and the system behaves in the opposite fashion as the form precedes function model, with the transcription network leading the space network. Between CP1 and CP2 in both models, the system fine tunes, or feedback between the networks leads to the optimum configuration for terminal differentiation at CP2.
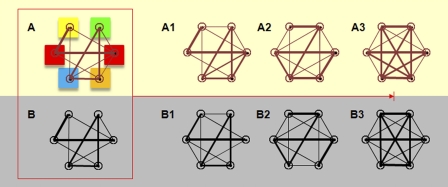
A network view of differentiation showing spatial and transcriptome networks in which nodes are genes or chromosomes. (A–B) Networks representing the signature of the undifferentiated state, in which some genes are connected, and some are not. The connectivity is weighted; that is, the strength of the connection depends on the gene pair, which is shown by edge thickness. In A, the colored boxes indicate their subnuclear location as described in Fig. 1, which defines node dynamics and, therefore, network edges (spatial distances). In B, the connectivity is also weighted, which defines gene coregulation. A1–A3 and B1–B3 represent the organization of the spatial and transcriptome networks during differentiation. The networks mutually influence each other, and fine tuning over time gives the differentiated system a unique network signature.
References
-
- Albiez H., Cremer M., Tiberi C., Vecchio L., Schermelleh L., Dittrich S., Küpper K., Joffe B., Thormeyer T., von Hase J., et al. 2006. Chromatin domains and the interchromatin compartment form structurally defined and functionally interacting nuclear networks. Chromosome Res. 14:707–733 10.1007/s10577-006-1086-x - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
