

A Nup133-dependent NPC-anchored network tethers centrosomes to the nuclear envelope in prophase
- PMID: 21383080
- PMCID: PMC3051818
- DOI: 10.1083/jcb.201007118
A Nup133-dependent NPC-anchored network tethers centrosomes to the nuclear envelope in prophase
Abstract
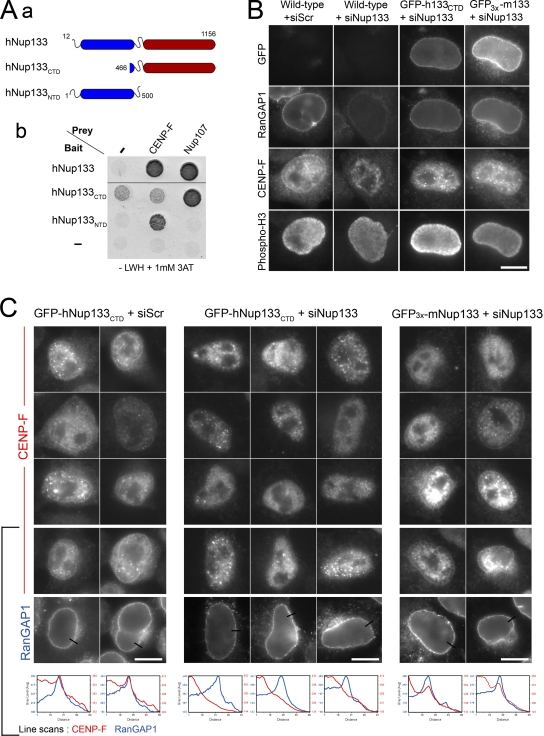
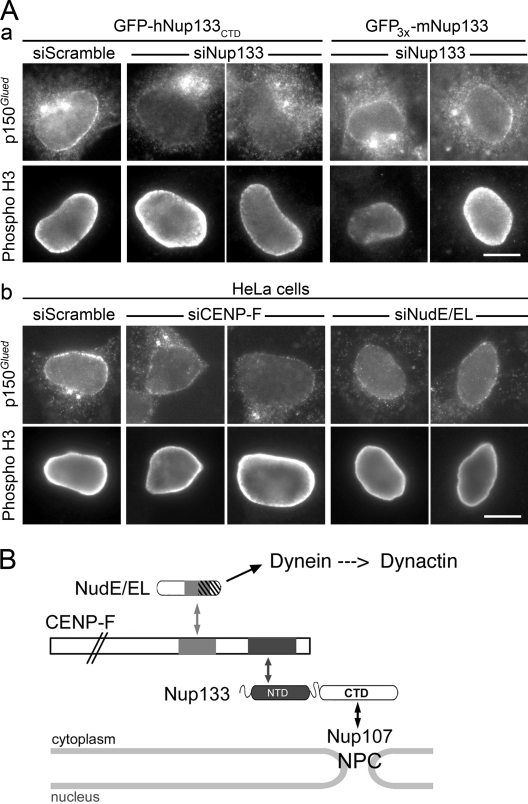
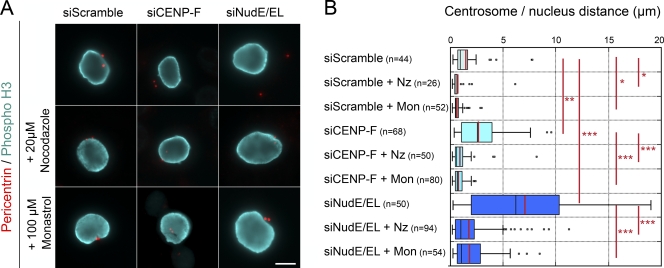
Centrosomes are closely associated with the nuclear envelope (NE) throughout the cell cycle and this association is maintained in prophase when they separate to establish the future mitotic spindle. At this stage, the kinetochore constituents CENP-F, NudE, NudEL, dynein, and dynactin accumulate at the NE. We demonstrate here that the N-terminal domain of the nuclear pore complex (NPC) protein Nup133, although largely dispensable for NPC assembly, is required for efficient anchoring of the dynein/dynactin complex to the NE in prophase. Nup133 exerts this function through an interaction network via CENP-F and NudE/EL. We show that this molecular chain is critical for maintaining centrosome association with the NE at mitotic entry and contributes to this process without interfering with the previously described RanBP2-BICD2-dependent pathway of centrosome anchoring. Finally, our study reveals that tethering of centrosomes to the nuclear surface at the G2/M transition contributes, along with other cellular mechanisms, to early stages of bipolar spindle assembly.
Figures






References
-
- Ban R., Matsuzaki H., Akashi T., Sakashita G., Taniguchi H., Park S.Y., Tanaka H., Furukawa K., Urano T. 2009. Mitotic regulation of the stability of microtubule plus-end tracking protein EB3 by ubiquitin ligase SIAH-1 and Aurora mitotic kinases. J. Biol. Chem. 284:28367–28381 10.1074/jbc.M109.000273 - DOI - PMC - PubMed
-
- Belgareh N., Rabut G., Baï S.W., van Overbeek M., Beaudouin J., Daigle N., Zatsepina O.V., Pasteau F., Labas V., Fromont-Racine M., et al. 2001. An evolutionarily conserved NPC subcomplex, which redistributes in part to kinetochores in mammalian cells. J. Cell Biol. 154:1147–1160 10.1083/jcb.200101081 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
