

Universal rule for the symmetric division of plant cells
- PMID: 21383128
- PMCID: PMC3076879
- DOI: 10.1073/pnas.1011866108
Universal rule for the symmetric division of plant cells
Abstract
The division of eukaryotic cells involves the assembly of complex cytoskeletal structures to exert the forces required for chromosome segregation and cytokinesis. In plants, empirical evidence suggests that tensional forces within the cytoskeleton cause cells to divide along the plane that minimizes the surface area of the cell plate (Errera's rule) while creating daughter cells of equal size. However, exceptions to Errera's rule cast doubt on whether a broadly applicable rule can be formulated for plant cell division. Here, we show that the selection of the plane of division involves a competition between alternative configurations whose geometries represent local area minima. We find that the probability of observing a particular division configuration increases inversely with its relative area according to an exponential probability distribution known as the Gibbs measure. Moreover, a comparison across land plants and their most recent algal ancestors confirms that the probability distribution is widely conserved and independent of cell shape and size. Using a maximum entropy formulation, we show that this empirical division rule is predicted by the dynamics of the tense cytoskeletal elements that lead to the positioning of the preprophase band. Based on the fact that the division plane is selected from the sole interaction of the cytoskeleton with cell shape, we posit that the new rule represents the default mechanism for plant cell division when internal or external cues are absent.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
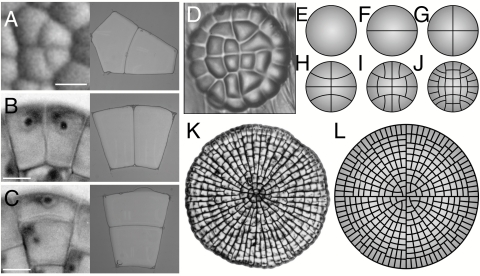
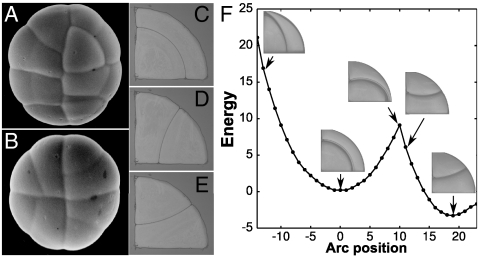
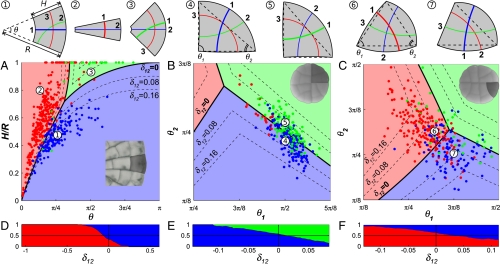
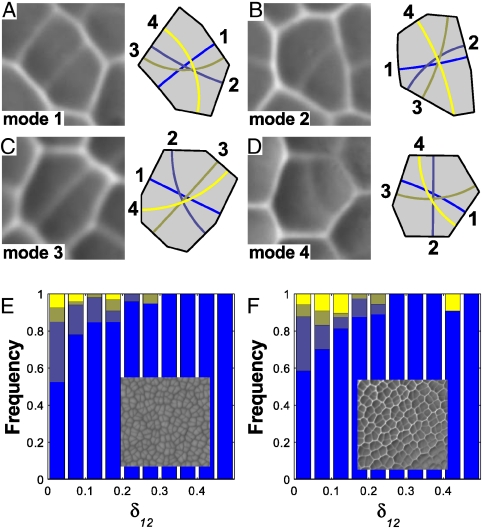
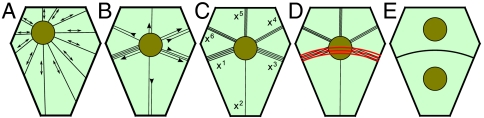
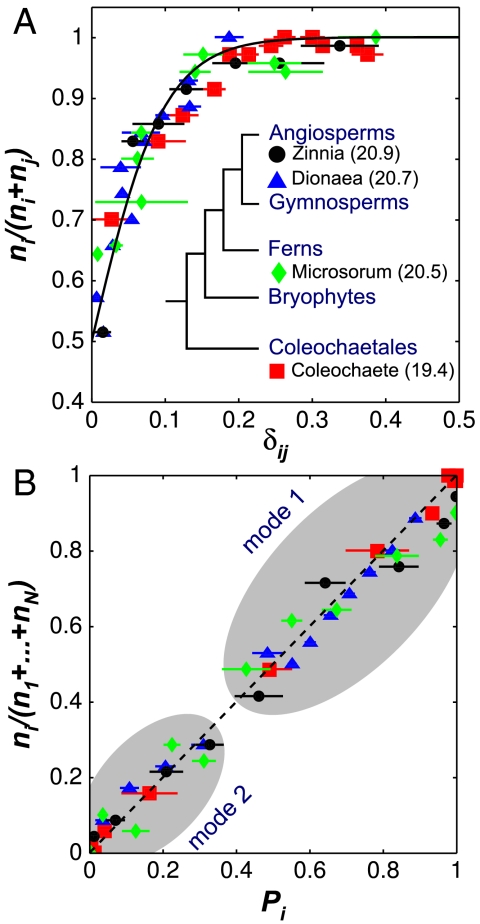
Comment in
-
Inherent randomness of cell division patterns.Proc Natl Acad Sci U S A. 2011 Apr 12;108(15):5933-4. doi: 10.1073/pnas.1103212108. Epub 2011 Apr 4. Proc Natl Acad Sci U S A. 2011. PMID: 21464313 Free PMC article. No abstract available.
References
-
- Bjerknes M. Physical theory of the orientation of astral mitotic spindles. Science. 1986;234:1413–1416. - PubMed
-
- Strauss B, Adams RJ, Papalopulu N. A default mechanism of spindle orientation based on cell shape is sufficient to generate cell fate diversity in polarised Xenopus blastomeres. Development. 2006;133:3883–3893. - PubMed
-
- Lintilhac PM, Vesecky TB. Stress-induced alignment of division plane in plant tissues grown in vitro. Nature. 1984;307:363–364.
-
- Yeoman MM, Brown R. Effects of mechanical stress on the plane of cell division in developing callus cultures. Ann Bot. 1971;35:1102–1112.
-
- Théry M, Jiménez-Dalmaroni A, Racine V, Bornens M, Jülicher F. Experimental and theoretical study of mitotic spindle orientation. Nature. 2007;447:493–496. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
