

Searching for simplicity in the analysis of neurons and behavior
- PMID: 21383186
- PMCID: PMC3176616
- DOI: 10.1073/pnas.1010868108
Searching for simplicity in the analysis of neurons and behavior
Abstract
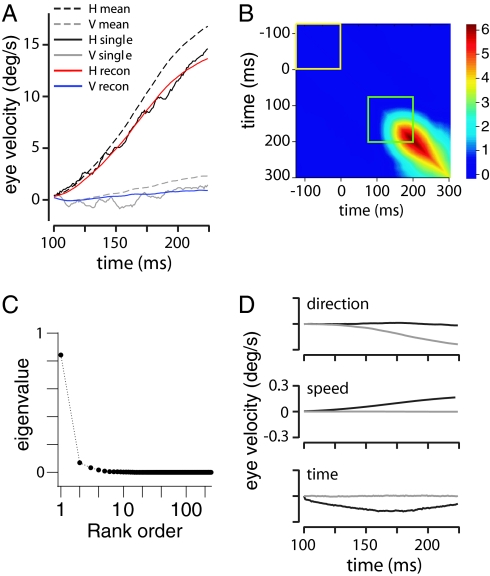
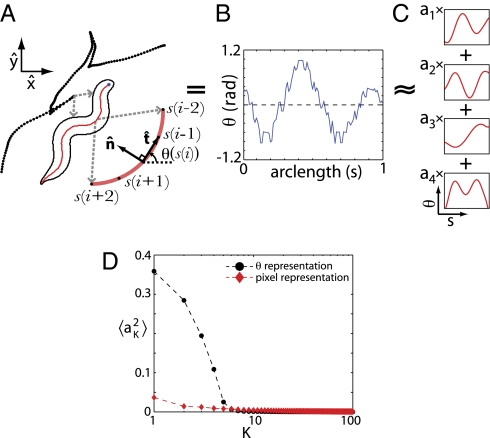
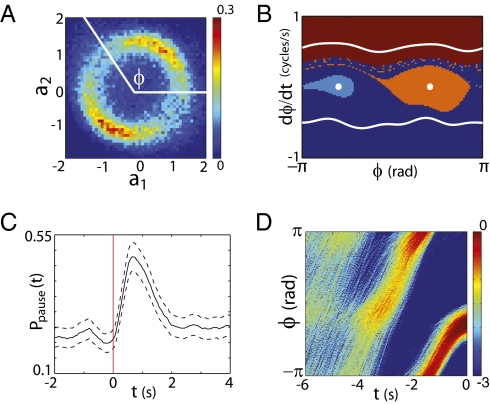
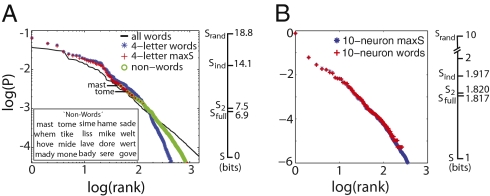
What fascinates us about animal behavior is its richness and complexity, but understanding behavior and its neural basis requires a simpler description. Traditionally, simplification has been imposed by training animals to engage in a limited set of behaviors, by hand scoring behaviors into discrete classes, or by limiting the sensory experience of the organism. An alternative is to ask whether we can search through the dynamics of natural behaviors to find explicit evidence that these behaviors are simpler than they might have been. We review two mathematical approaches to simplification, dimensionality reduction and the maximum entropy method, and we draw on examples from different levels of biological organization, from the crawling behavior of Caenorhabditis elegans to the control of smooth pursuit eye movements in primates, and from the coding of natural scenes by networks of neurons in the retina to the rules of English spelling. In each case, we argue that the explicit search for simplicity uncovers new and unexpected features of the biological system and that the evidence for simplification gives us a language with which to phrase new questions for the next generation of experiments. The fact that similar mathematical structures succeed in taming the complexity of very different biological systems hints that there is something more general to be discovered.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nelson WL. Physical principles for economies of skilled movements. Biol Cybern. 1983;46:135–147. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
