

Reciprocal and activity-dependent regulation of surface AMPA and NMDA receptors in cultured neurons
- PMID: 21383896
- PMCID: PMC3047273
Reciprocal and activity-dependent regulation of surface AMPA and NMDA receptors in cultured neurons
Abstract
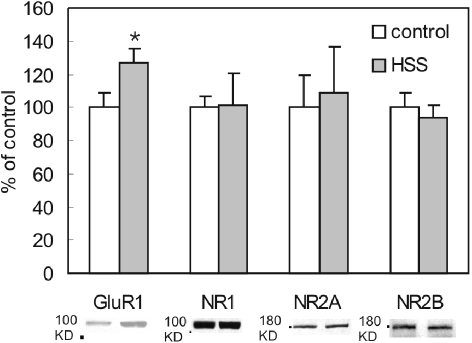
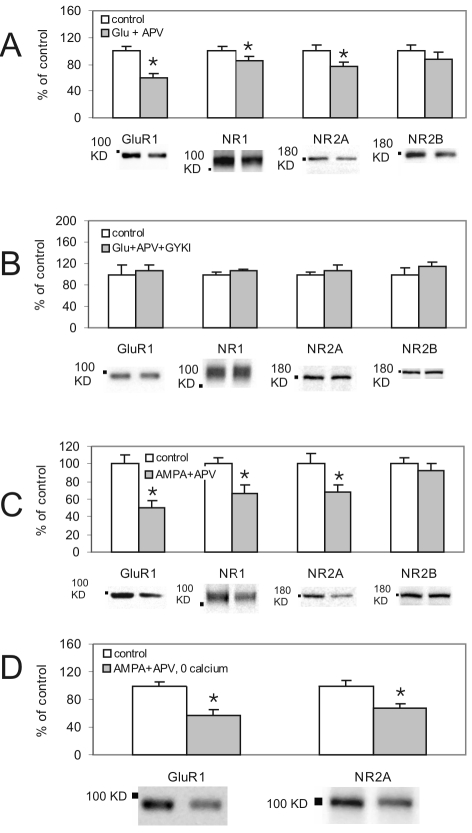
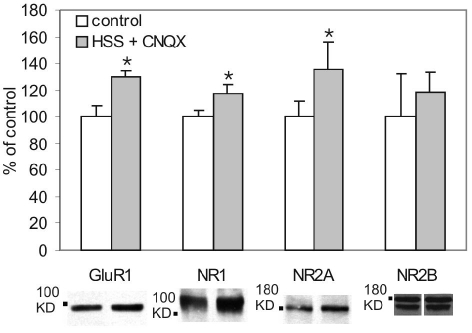
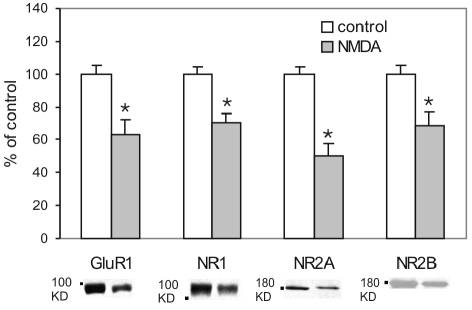
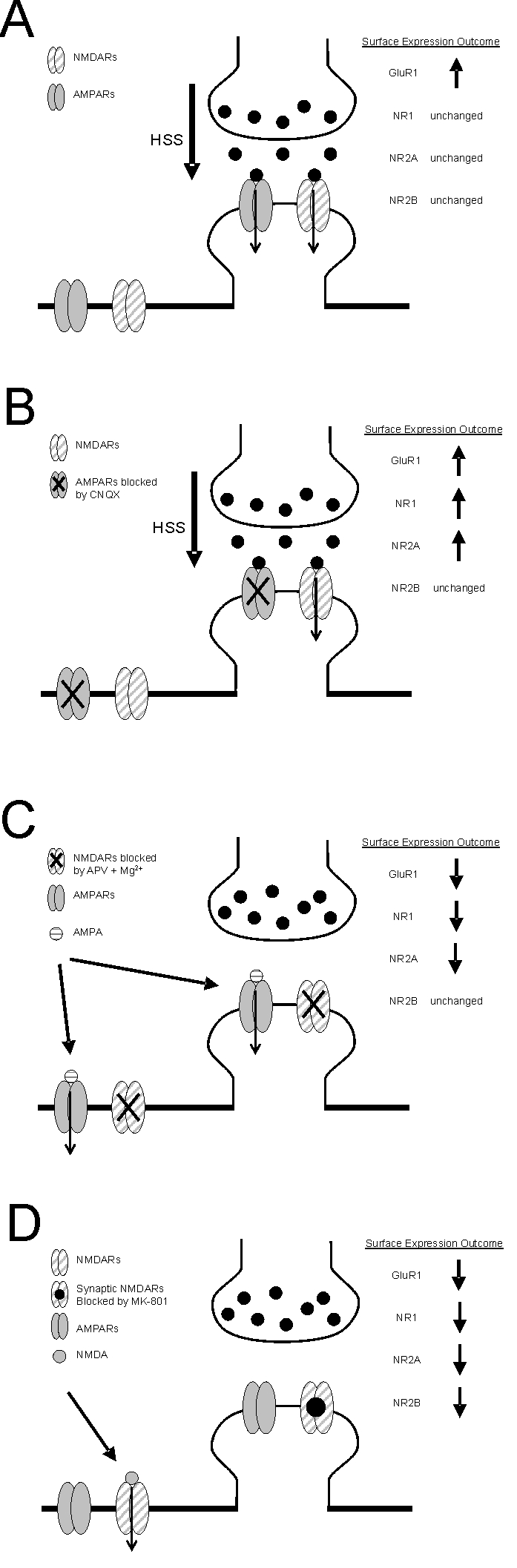
Activation of NMDA receptors (NMDARs) can modulate excitatory synaptic transmission in the central nervous system by dynamically altering the number of synaptic AMPA receptors (AMPARs). The surface expression of NMDARs themselves is also subject to modulation in an activity-dependent manner. In addition to NMDAR-induced changes in AMPAR expression, AMPARs have also been found to regulate their own surface expression, independently of NMDARs. However, whether or not AMPARs and NMDARs might reciprocally regulate their surface expression has not previously been systematically explored. We utilized surface biotinylation assays and stimulation protocols intended to selectively stimulate various glutamate receptor subpopulations (e.g. AMPARs vs NMDARs; synaptic vs extrasynaptic). We reveal that activation of synaptic NMDARs increases the surface expression of both NMDAR and AMPAR subunits, while activation of extrasynaptic NMDAR produces the opposite effect. Surprisingly, we find that selective activation of AMPARs reduces the surface expression of not only AMPARs but also of NMDARs. These results suggest that both AMPARs and NMDARs at synaptic sites are subject to modulation by multiple signalling pathways in an activity-dependent way.
Figures
References
-
- Derkach VA, Oh MC, Guire ES, Soderling TR. Regulatory mechanisms of AMPA receptors in synaptic plasticity. Nat Rev Neurosci. 2007;8:101–113. - PubMed
-
- Groc L, Choquet D. AMPA and NMDA glutamate receptor trafficking: multiple roads for reaching and leaving the synapse. Cell Tissue Res. 2006;326:423–438. - PubMed
-
- Malinow R, Malenka RC. AMPA receptor trafficking and synaptic plasticity. Annu Rev Neurosci. 2002;25:103–126. - PubMed
LinkOut - more resources
Full Text Sources