

Deciphering the nuclear bile acid receptor FXR paradigm
- PMID: 21383957
- PMCID: PMC3049226
- DOI: 10.1621/nrs.08005
Deciphering the nuclear bile acid receptor FXR paradigm
Abstract
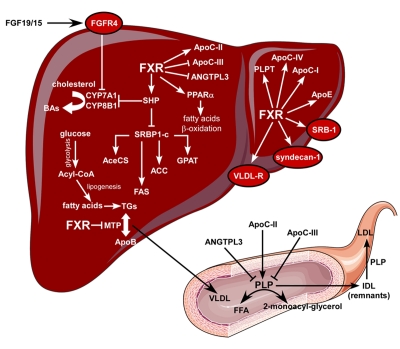
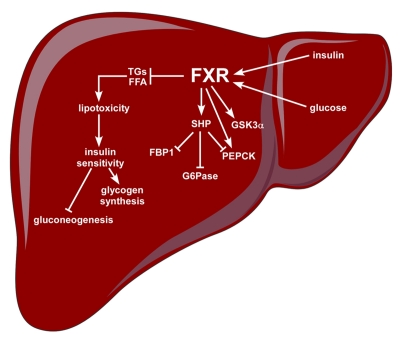
Originally called retinoid X receptor interacting protein 14 (RIP14), the farnesoid X receptor (FXR) was renamed after the ability of its rat form to bind supra-physiological concentrations of farnesol. In 1999 FXR was de-orphanized since primary bile acids were identified as natural ligands. Strongly expressed in the liver and intestine, FXR has been shown to be the master transcriptional regulator of several entero-hepatic metabolic pathways with relevance to the pathophysiology of conditions such as cholestasis, fatty liver disease, cholesterol gallstone disease, intestinal inflammation and tumors. Furthermore, given the importance of FXR in the gut-liver axis feedbacks regulating lipid and glucose homeostasis, FXR modulation appears to have great input in diseases such as metabolic syndrome and diabetes. Exciting results from several cellular and animal models have provided the impetus to develop synthetic FXR ligands as novel pharmacological agents. Fourteen years from its discovery, FXR has gone from bench to bedside; a novel nuclear receptor ligand is going into clinical use.
Figures
References
-
- Ananthanarayanan M., Li S., Balasubramaniyan N., Suchy F. J., Walsh M. J. Ligand-dependent activation of the farnesoid X-receptor directs arginine methylation of histone H3 by CARM1. J Biol Chem. 2004;279:54348–57. - PubMed
-
- Anisfeld A. M., Kast-Woelbern H. R., Lee H., Zhang Y., Lee F. Y., Edwards P. A. Activation of the nuclear receptor FXR induces fibrinogen expression: a new role for bile acid signaling. J Lipid Res. 2005;46:458–68. - PubMed
-
- Anisfeld A. M., Kast-Woelbern H. R., Meyer M. E., Jones S. A., Zhang Y., Williams K. J., Willson T., Edwards P. A. Syndecan-1 expression is regulated in an isoform-specific manner by the farnesoid-X receptor. J Biol Chem. 2003;278:20420–8. - PubMed
-
- Araya Z., Wikvall K. 6alpha-hydroxylation of taurochenodeoxycholic acid and lithocholic acid by CYP3A4 in human liver microsomes. Biochim Biophys Acta. 1999;1438:47–54. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
